

Bxb1たちはどう回るか
―DNA組換え酵素Bxb1の回転運動の可視化に成功!―
- プレスリリース
2026年6月9日
東京大学
発表のポイント
- ラージセリンリコンビナーゼBxb1は、特定の塩基配列(attPとattB)を持つDNAの間の組換え反応を触媒するため、ゲノム編集技術に利用されている。
- クライオ電子顕微鏡を用いて、DNA組換え反応中の4つの状態にあるBxb1–attP–attB複合体の立体構造を決定した。
- attPとattBがBxb1によって切断された後、複合体の中心に存在する疎水的で平坦な表面を介して左右の二量体(Bxb1–attP断片とBxb1–attB断片)が約180度回転し、DNA鎖交換が進行する様子を可視化した。
- 構造情報に基づきBxb1を改変し、認識配列の異なる変異体や組換え活性の向上した変異体を作製した。
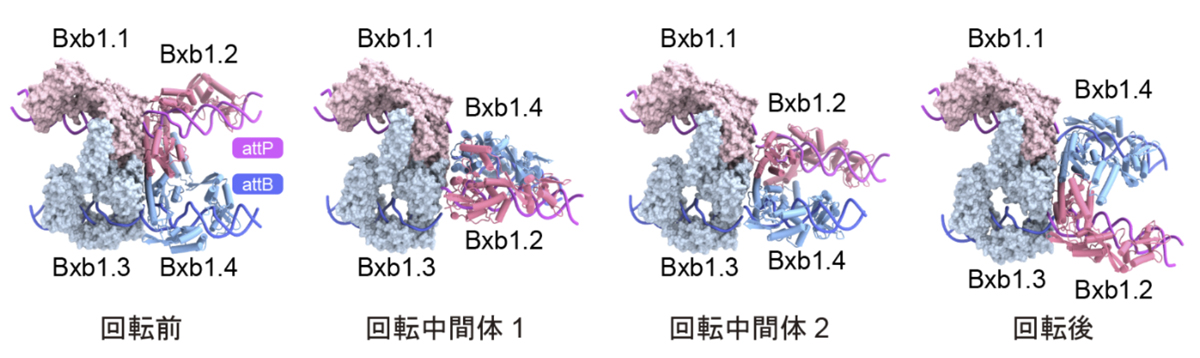
概要
ラージセリンリコンビナーゼ(注1)はDNA組換え活性を持つ酵素であり、近年、長鎖DNAをゲノム中の狙った位置に挿入するゲノム編集技術への応用において注目されています。その一つであるBxb1はattP配列とattB配列(注2)の間のDNA組換え反応を触媒しますが、2種類のDNA配列に結合し、DNAを特定の位置で切断し、DNA断片を交換した後、それらを再結合するという複数のステップからなる複雑な反応メカニズムには不明な点が残されていました。今回、東京大学大学院工学系研究科化学生命工学専攻の相馬哲兵大学院生、平泉将浩助教、同大学先端科学技術センターの西増弘志教授らのグループは、Harvard Medical SchoolのChristopher W. Fell研究員、Jonathan S. Gootenberg主任研究員、Omar O. Abudayyeh主任研究員らのグループとの共同研究として、クライオ電子顕微鏡(注3)を用いて、4つの異なる状態にあるBxb1-attP-attB複合体の立体構造を決定しました。その結果、2分子のBxb1がattPとattBに異なる様式で結合し、非対称な四量体の複合体を形成することが明らかになりました。さらに、DNA切断後に左右の二量体が約180度回転し、DNA鎖交換が進行する様子の可視化に成功しました。さらに、研究チームは得られた構造情報に基づいてBxb1を改変し、DNA選択性の異なる変異体や、組換え活性の向上した変異体の作製にも成功しました。本研究成果は、ラージセリンリコンビナーゼの作動原理の理解を深めるとともに、長鎖DNAの標的部位への挿入を可能にする次世代ゲノム編集技術の開発基盤となることが期待されます。本成果は、2026年6月8日に「Molecular Cell」オンライン版に掲載されました。
ラージセリンリコンビナーゼは1990年代から研究が進み、Bxb1についても2000年代にはDNA組換え反応の基本的な性質が明らかにされていましたが、複合体構造は長く不明のままでした。今回、構造解析技術の進歩に加えて、反応中間体を捉えるための工夫によって、その複合体構造を明らかにできた過程は、今後の研究人生でも忘れられない経験になると思います。さらに予想外だったのは、回転の中間状態に対応する構造まで得られたことです。構造解析の途中で、当初は除いていた粒子の中に回転中間体が含まれていることに気づいたときの衝撃は非常に大きく、今でも強く印象に残っています。今回、自分たちが明らかにした構造を出発点として、Bxb1のゲノム編集への応用研究がさらに進むことを期待しています。
(東京大学大学院工学系研究科 相馬哲兵 大学院生)
ラージセリンリコンビナーゼ(LSR)は、ゲノム編集への応用面で注目されているだけでなく、(1)似て非なる塩基配列(attPとattB)を持つ2分子の二本鎖DNAとの結合、(2)4本のDNA鎖の特定の位置での切断および5′-ホスホセリン共有結合中間体の形成、(3)左右二量体の回転を伴うDNA鎖の交換、(4)DNA鎖の再結合という複数のステップからなるユニークなメカニズムでDNA組換え反応を触媒する基礎的にも非常に興味深いタンパク質です。LSRは1990年代から研究されており、生化学的な研究からDNA組換えメカニズムが提唱されていましたが、複合体分子の柔軟性のため構造解析が困難であり、DNAと結合した機能的な複合体の立体構造は不明でした。特に、分子の大規模な回転は他の酵素反応では見られないユニークなメカニズムですが、どのように回転しているかは大きな謎でした。
2020年に研究室を立ち上げ、2022年に最初の4年生として配属された相馬くんがスタッフの指導の下、クライオ電子顕微鏡構造解析に精力的に取り組んでくれた結果、2023年にBxb1-attP-attB複合体の立体構造を決定することができました。予想外にも、回転反応の途中に対応する2つの中間体構造も明らかになり、Bxb1が疎水的な(ヌルヌルした)平らな分子表面を利用して実際に回転する様子の可視化に世界で初めて成功しました。この研究成果は、30年来謎に包まれていたユニークなDNA組換えメカニズムを明らかにし、ゲノム編集技術の高度化にも直結する重要な発見だったため、2024年7月にScience誌に投稿しました。10月にレビューコメントが返ってきました。4人のレビュアーのうち1名は「Overall, this is an excellent study, but probably a better fit for another journal.」としてScience誌への掲載に否定的でしたが、他の3名は「Overall, this work is impactful and well performed.」、「Collectively, this work visualized the recombination process and lays a solid foundation for engineering Bxb1 into a tool for genome integration.」といった意見で、研究成果の重要性を評価してくれました。レビューコメントに従い追加実験を行い、2025年1月にアクセプトを期待してリバイズ原稿を投稿しましたが、2か月後に返ってきた結果は、予想外にもリジェクトでした。最初は否定的だったレビュアーは出版を支持してくれた一方、当初好意的だったレビュアーの1人が「Bxb1改変体のわずかな活性向上(野生型の1.5倍)と誤解を招く主張を踏まえると、この研究の出版(ましてやScience誌での掲載)を支持することはできません。高品質な構造解析があるだけに残念です。」というコメントが原因でした。誤解があったため反論しましたが、残念ながら結果は変わりませんでした。
その後、Nature誌とCell誌に投稿しましたが、査読には回りませんでした。失意の中、Molecular Cell誌に投稿したところ、査読に回った後、6か月間レスポンスがありませんでした。埒が明かないため、取り下げの可能性にも言及して4度目の問い合わせメールを送ったところ、ようやく返信が来ました。2人のレビュアーのうち1人が否定的だったため(本質的には好意的でしたが)、エディターが3人目のレビュアーを追加したことが、長い時間がかかった要因でした。幸いにも、3人目のレビュアーは非常に好意的でした。その後、適切な追加実験を行い、リバイズ原稿を投稿し、2026年5月についに論文がアクセプトされました。Science誌に投稿してから2年近くが経過していました。
アクセプトまでには多くの困難がありましたが、複数のジャーナルでのリバイズを経る中で、データの量・質および文章が大きく改善され、最終的にはとてもよい論文になりました。最近、2つの競合グループから(回転中間体は含まれていませんが)LSR-DNA複合体構造に関するプレプリントが発表されましたが、最初の論文として発表できたことをうれしく思います。最後に、丁寧に査読してくれた多くのレビュアーの方々に感謝いたします。
(東京大学先端科学技術研究センター 西増弘志 教授)
発表内容
ラージセリンリコンビナーゼは、ファージの持つDNA組換え酵素であり、ファージゲノムの宿主ゲノムへの組み込みや切り出しに関与します。Bxb1はその一種であり、attPとattBと呼ばれる似て非なるDNA配列の間の組換え反応を触媒します。Bxb1はヒト細胞においても機能することから、近年、長鎖DNAを狙った場所に挿入するゲノム編集(注4)に利用されています。特に、最新のゲノム編集技術であるPASTEやPASSIGEは、プライムエディター(注5)を用いてゲノム中にattB配列を導入した後、attP配列をもつドナーDNAをBxb1によって組み込む技術であり、長鎖DNAを標的部位に導入できるゲノム編集技術として注目されています。生化学的解析から、Bxb1は二量体としてattPとattBにそれぞれ結合した後、四量体の複合体を形成し、2つのDNAを特定の位置で切断した後、左右の2つの二量体が約180度回転することによりDNA鎖が交換されることが示唆されていました(図1)。しかし、Bxb1がattPとattBに結合し、DNA鎖の切断、交換、再結合を段階的に触媒する詳細な仕組みは、1990年代のラージセリンリコンビナーゼの発見以来30年以上、謎に包まれていました。

今回、本研究チームはクライオ電子顕微鏡解析を行い、DNA組換え反応中の異なる段階に対応する4つの状態にあるBxb1-attP-attB複合体の立体構造を決定しました(図2)。その結果、4分子のBxb1が1分子のattPと1分子のattBに結合し、非対称な四量体の複合体を形成することが明らかになりました。
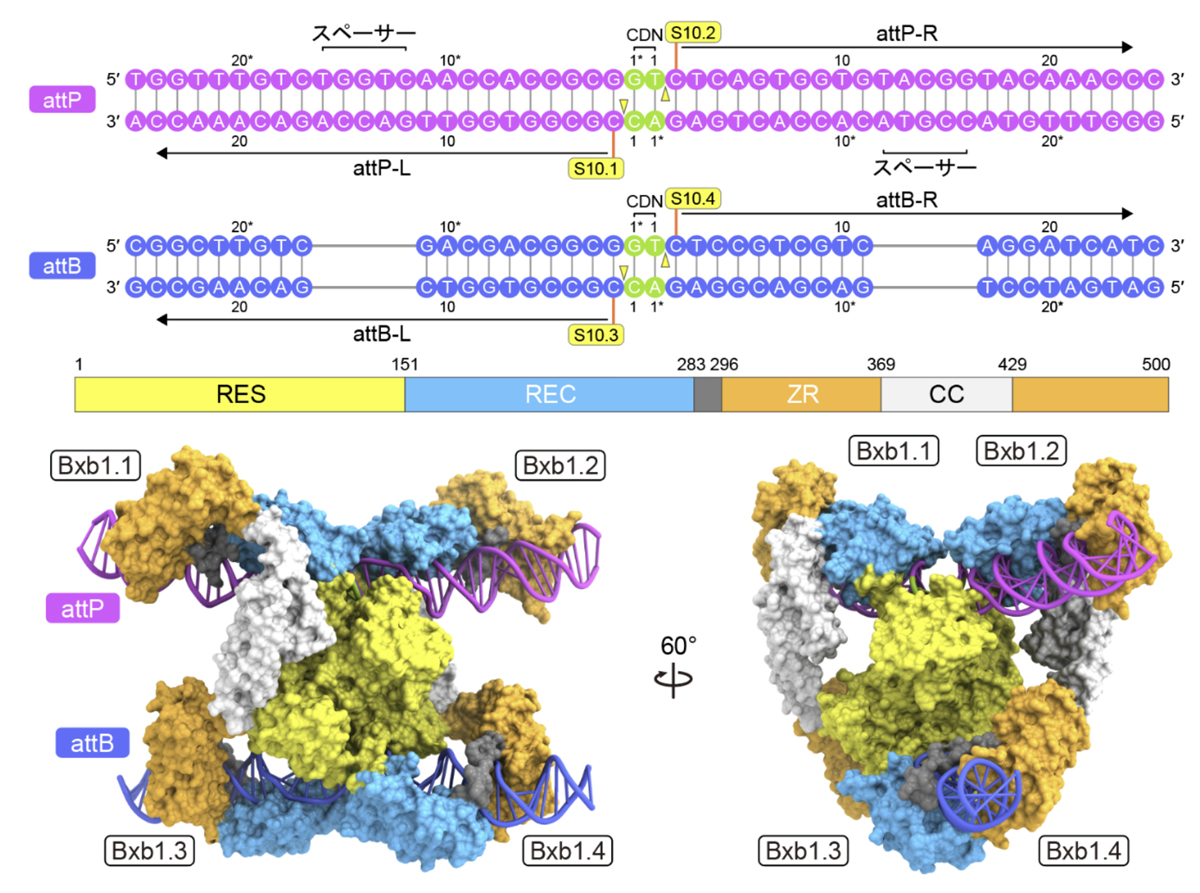
今回の構造から、Bxb1がattPとattBの塩基配列をどのように認識しているかも明らかになりました。中心側のDNA領域はBxb1のRESドメインとRECドメインによって認識される一方、外側のDNA領域はZRドメインによって認識されていました(図3)。注目すべきことに、attPに結合したBxb1とattBに結合したBxb1ではZRドメインの位置が異なり、この違いによってCCドメイン同士の非対称な相互作用が生じることがわかりました。CCドメインを削除するとBxb1のDNA組換え活性が消失したことから、CCドメインを介した相互作用が機能的な四量体の形成に必須であることが明らかになりました。

(下)attP結合型Bxb1とattB結合型Bxb1の構造比較。スペーサー配列の有無により、ZRドメインとCCドメインの配置がattP結合型Bxb1とattB結合型Bxb1で異なる。
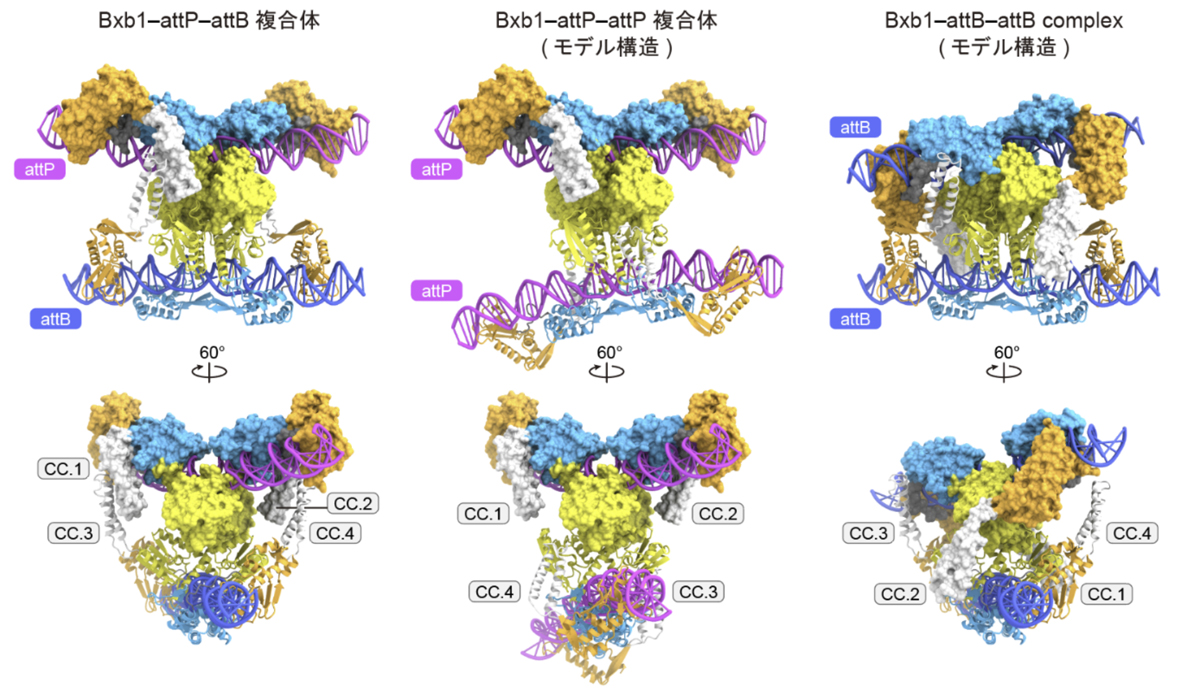
attP-attBに結合した四量体の複合体構造と、attP-attPおよびattB-attBに結合した複合体の予測構造の比較から、Bxb1がattPのみ、または、attBのみと結合した場合、CCドメイン同士の相互作用が形成されず、機能的な四量体が形成されないことが示唆されました(図4)。これらの構造比較から、Bxb1がattP-attPやattB-attBの間の組換え反応は触媒せず、attP-attBの間の組換え反応を選択的に触媒する仕組みが明らかになりました。
今回決定した4つの構造から、DNA鎖交換の過程で左右の2つの二量体が回転する様子も可視化されました(図5)。Bxb1は、attPとattBを特定の位置で切断し共有結合中間体を形成した後、複合体の中央に存在する疎水性で平坦な表面を介して、一方の二量体が他方に対して約180度回転し、その結果、DNA鎖が交換され、最終的に再結合が起こることでDNAの組換えが完了します。構造比較の結果、回転中間体1、回転中間体2、回転後の状態は、回転前の状態に対して左右の二量体がそれぞれ約120度、-40度、160度回転した位置に存在することがわかりました。また、これらの回転中間体の構造から、2つの二量体が解離することなく回転し、DNA組換え反応を触媒することが示唆されました。
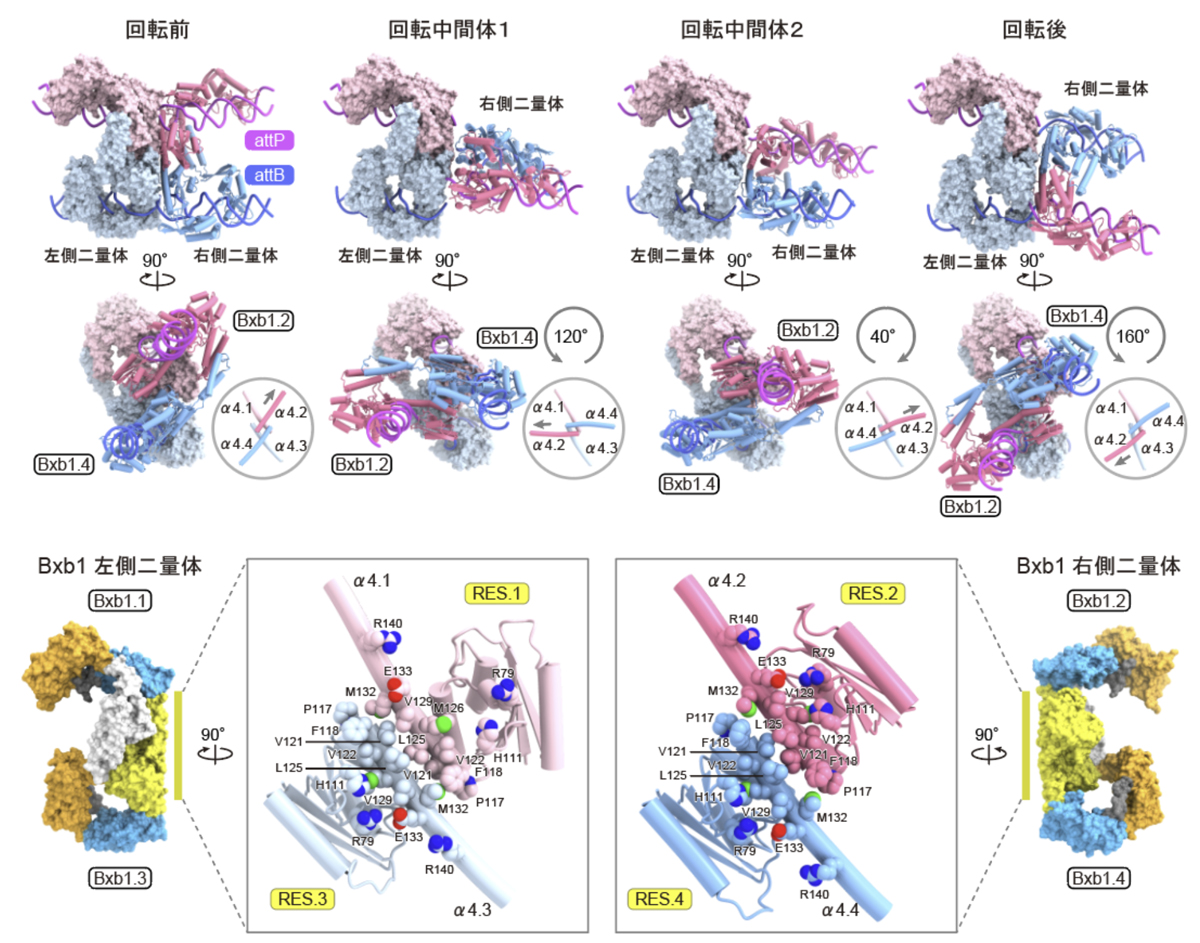
(下)疎水性界面の拡大図。RESドメインが形成する界面は主に疎水性残基から構成され平坦な形状をしているため、左右の二量体の間の回転が可能である。
研究チームは、今回得られた構造情報をもとに、Bxb1の機能改変にも取り組みました。まず、DNA塩基と相互作用するアミノ酸残基に変異を導入することにより、Bxb1のDNA配列選択性が変化するかを検討しました。ヒト培養細胞における変異体解析の結果、N141やR237などの残基を別のアミノ酸に置換すると、Bxb1のattP/attB配列に対する選択性が変化することが明らかになりました(図6)。さらに、DNAのリン酸骨格近傍に位置する複数の残基を正電荷を持つリジン残基に置換することにより、DNAとの相互作用を強化し、野生型より高い組換え活性を示すBxb1変異体を作製することにも成功しました。特に、V12K/Q226K/H334K変異を持つenBxb1、および、enBxb1とプライムエディターを組み合わせたenPASTEでは、複数の標的部位においてDNA組み込み効率が平均して約1.5倍向上することがわかりました。
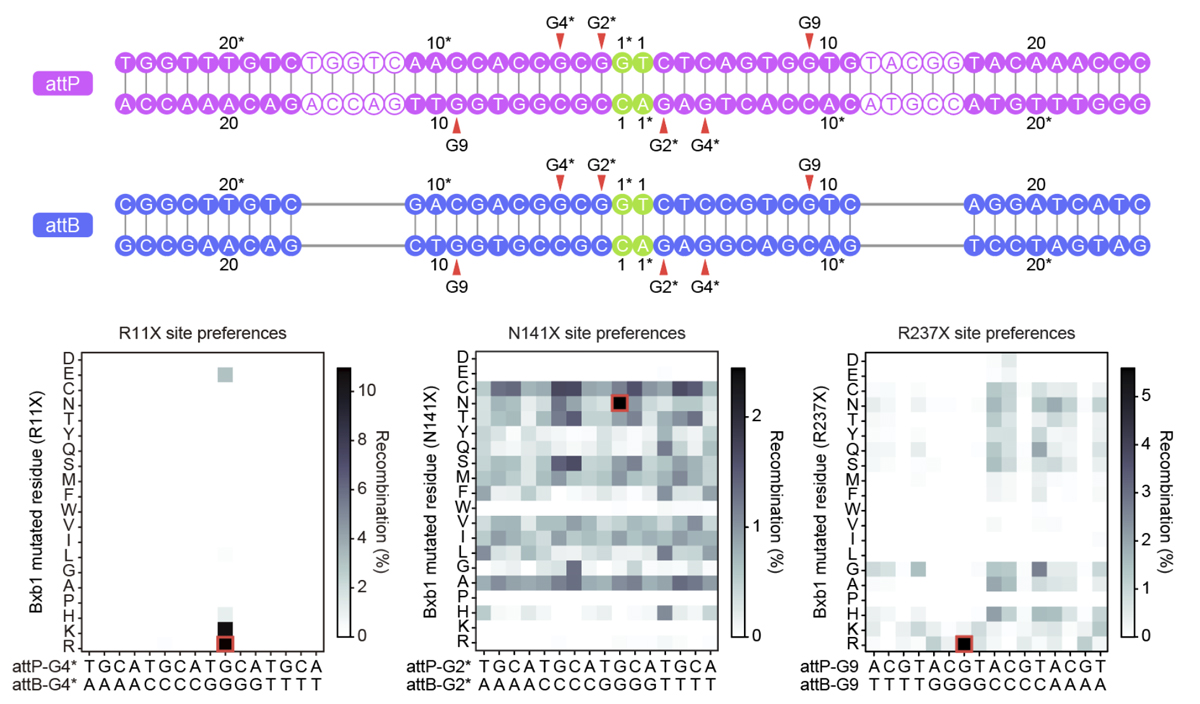
(下)R11X、N141X、R237X変異体のDNA配列選択性の評価。ヒト培養細胞に各Bxb1変異体と、変異型attP/attBプラスミドの組み合わせを導入し、組換え活性を測定した。ヒートマップは各変異体の組換え効率を示す。赤枠は野生型Bxb1と野生型attP/attB配列の組み合わせを示す。R11の置換のほとんどが活性を消失させた一方、N141およびR237の置換では、配列選択性の変化が観察された。
本研究により、Bxb1がattPとattBの両方を認識して非対称な複合体を形成し、二量体の回転によってDNA組換えが進行する分子メカニズムが明らかになりました。さらに、構造情報を利用することで、標的配列選択性や活性の変化したBxb1改変体を設計できることも示されました。これらの成果は、ラージセリンリコンビナーゼの作動原理の理解を深めるだけでなく、長鎖DNAの組み込みを可能にするゲノム編集技術の改良につながることが期待されます。
発表者・研究者等情報
東京大学
大学院工学系研究科
相馬 哲兵 大学院生
平泉 将浩 助教
先端科学技術研究センター
山下 恵太郎 准教授
西増 弘志 教授
兼:東京大学大学院工学系研究科 教授
Harvard Medical School
Christopher W. Fell Postdoctoral Research Fellow
Jonathan S. Gootenberg Investigator
Omar O. Abudayyeh Investigator
論文情報
- 雑誌名:
- 「Molecular Cell」(オンライン版:6月8日)
- 題 名:
- Structure and engineering of the large serine recombinase Bxb1 for gene integration
- 著者名:
- Teppei Soma†, Masahiro Hiraizumi†, Christopher W. Fell†, Dario Tagliaferri, Jason Lequyer, Sae Okazaki, Yukari Isayama, Kazuki Kato, Sworaj Sapkota, Harsh Ramani, Benjamin Arya, Cian Schmitt-Ulms, Keitaro Yamashita, Jonathan S. Gootenberg*, Omar O. Abudayyeh*, Hiroshi Nishimasu*(†共同筆頭著者、*責任著者)
- DOI:
- https://doi.org/10.1016/j.molcel.2026.05.018
研究助成
本研究は、日本学術振興会(JSPS)科研費「特別研究員奨励費(課題番号:25KJ0809)」、「若手研究(課題番号:23K14133)」、「学術変革領域研究(A)(課題番号:21H05281)」、「基盤研究(A)(課題番号:22H00403)」、「基盤研究(S)(課題番号:25H00436)」、国立研究開発法人日本医療研究開発機構(AMED)「創薬等先端技術支援プラットフォーム(BINDS)(課題番号:JP21am0101115、支援番号:6464)」、科学技術振興機構(JST)戦略的創造研究推進事業「ACT-X(課題番号:JPMJAX232F)」、「CREST(課題番号:JPMJCR23B6)」、武田科学振興財団「武田報彰医学研究助成」、稲盛財団InaRISフェローシップなどの支援により実施されました。
用語解説
- (注1)ラージセリンリコンビナーゼ
DNA組換え反応を触媒する酵素群の一つ。ファージが自身のDNAを宿主ゲノムに組み込む際に利用される。 - (注2)attP/attB配列
Bxb1が認識する2種類のDNA配列。attP(phage attachment site)はファージ側、attB(bacterial attachment site)は宿主側の配列に由来し、Bxb1はこの2つの配列間のDNA組換え反応を触媒する。 - (注3)クライオ電子顕微鏡
液体窒素冷却下でタンパク質などの分子に電子線を照射し、試料の観察を行うための装置。タンパク質や核酸の立体構造の決定に利用される。 - (注4)ゲノム編集
生物のもつ遺伝情報であるゲノムDNAの特定の塩基配列を、狙った位置で改変する技術。特定の塩基を置き換えたり、塩基配列を挿入したり削除したりすることができる。 - (注5)プライムエディター
Cas9ニッカーゼと逆転写酵素の融合タンパク質であり、ゲノム中の標的部位に塩基置換や挿入・欠失を導入することができる。
問合せ先
東京大学先端科学技術研究センター 構造生命科学分野
教授 西増 弘志(にします ひろし)
関連タグ
