

CRISPR-Cas9の分子進化を可視化!
- 研究成果
2026年1月30日
東京大学
発表のポイント
- ゲノム編集に利用されているCas9は、トランスポゾンの転移に関与するIscBから進化したことが知られていたが、その詳細は不明だった。
- クライオ電子顕微鏡を用いて、IscBからCas9への分子進化の中間体と考えられる2種類のIscBおよび2種類のCas9の立体構造を決定した。
- これら4つの立体構造と既知のIscBやCas9の立体構造の比較から、IscBからCas9への分子進化の過程において、RNAの小型化とタンパク質の大型化が起こり、DNAに対する特異性が向上したことが示唆された。
- 本研究成果は、高精度なゲノム編集ツールの設計や改良への応用が期待される。
概要
原核生物のCRISPR-Cas獲得免疫機構(注1)に関与するCas9タンパク質は、ガイドRNA(注2)と複合体を形成し、標的となる2本鎖DNAを切断します。近年、トランスポゾン(注3)の転移に関与するIscBタンパク質が、Cas9と同様にガイドRNAと協働してDNAを切断することが明らかとなりました。Cas9とIscBはゲノムDNAを狙った位置で切断できるため、ゲノム編集技術(注4)に応用されており、Cas9の機能を明らかにした研究者は2020年にノーベル化学賞を受賞しました。Cas9はIscBから進化してきたことが示唆されていましたが、その詳細は不明でした。今回、東京大学大学院工学系研究科化学生命工学専攻の長畑直人大学院生、山田崇太大学院生、先端科学技術研究センターの加藤一希講師(現東京科学大学)、西増弘志教授らの研究チームは、クライオ電子顕微鏡(注5)を用いて系統的に多様な2種類のIscB、および、2種類のCas9の立体構造を決定しました。これらの立体構造と生化学解析の結果、IscBからCas9への分子進化の過程において、RNAの小型化とタンパク質の大型化が起こり、DNAに対する特異性が向上したことが示唆されました。本研究成果は、高い特異性をもつ小型のゲノム編集ツールの開発への応用が期待されます。本成果は、2026年1月30日(英国時間)に英国科学誌「Nature Structural & Molecular Biology」オンライン版に掲載されました。
CRISPR-Cas9を利用した正確かつ簡便なゲノム編集技術は、農学、遺伝子治療、創薬など様々な分野の生命科学研究を大きく発展させてきました。今回の研究では、Cas9が分子進化の過程で、DNAを正確に切断する能力をどのように獲得してきたのか、その仕組みを推定することができました。一方で、分子進化の視点からCas9やその祖先であるIscBを眺めてみると、なお面白い謎が残されていることにも気付かされます。IscBのような祖先型のRNA誘導型システムはどのように誕生したのでしょうか。また、現存するIscBやCas9は今後どのように進化し、どのような機能を獲得していくのでしょうか。まだまだ興味は尽きません。(東京大学大学院工学系研究科 大学院生 長畑直人)
構造を見比べている中で、特異性が段階的に向上していることに気付いた時の感動は忘れられません。研究室に所属した当初から取り組んでいた研究を論文として発表することができ、感慨深く思います!(東京大学大学院工学系研究科 大学院生 山田崇太)
このプロジェクトを開始してから論文として発表するまでに5年かかりましたが、クライオ電子顕微鏡を用いて4つの複合体構造を決定し、CRISPR-Cas9の分子進化の詳細を世界にさきがけて解明できたことをうれしく思います。通常の論文に比べてデータ量が多く、論文としてまとめるのに苦労しましたが、長畑くんが中心となってがんばってくれて、美しい論文として発表することができました。ただ、Cell誌の査読において、Reviewer 1は非常に好意的だった一方、Reviewer 2は「Comparative structural analysis does not inform evolutionary history」と批判的であったため、リジェクトされたのは残念でした。「立体構造の比較から分子進化に関する知見は得られない」ということはないと思いますが、Cell誌に投稿した原稿には論理の飛躍があったのは確かでした。そこで、Reviewer 2のコメントを反映した原稿をNSMB誌に投稿し、最終的により洗練された論文として発表することができました。Reviewer 2、ありがとう!
(東京大学先端科学技術研究センター 教授 西増弘志)
発表内容
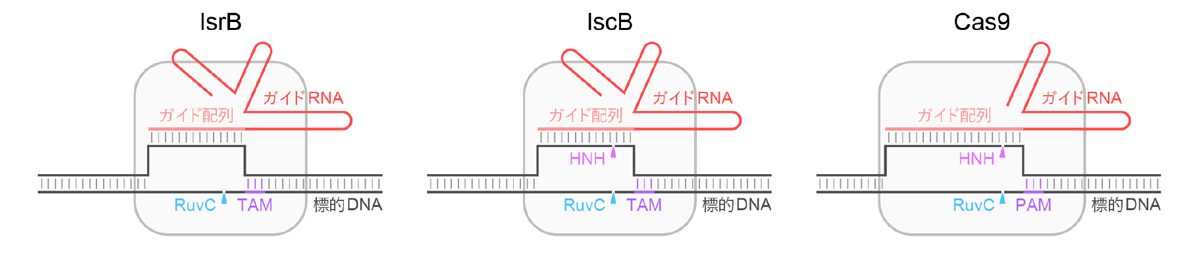
CRISPR-Cas9は原核生物のもつ獲得免疫機構であり、細胞内に侵入したウイルスなどの外来核酸を認識・切断することで生体防御を担っています。CRISPR配列には過去に感染した外来核酸の配列情報の一部が保存されており、そこからガイドRNAが産生されます。Cas9タンパク質はガイドRNAと複合体を形成し、RuvCおよびHNHヌクレアーゼドメイン(注6)を用いて、ガイドRNAの一部(ガイド配列)との塩基相補性およびPAM配列(注7)をもつ標的DNAを切断します(図1)。近年、トランスポゾンに関与するIsrBおよびIscBタンパク質が、Cas9と同様にガイドRNA依存的にTAM配列(注7)を含む標的DNAを切断することが明らかになりました(図1)。系統解析(注8)から、Cas9はIsrBからIscBを経て進化した可能性が示唆されていましたが、Cas9、IscB、およびIsrBはタンパク質とガイドRNAのサイズが大きく異なるため、分子進化の詳細は不明でした。本研究では、IscBからCas9への分子進化に着目し、その進化過程で生じた分子構造の変化を詳細に解析しました。
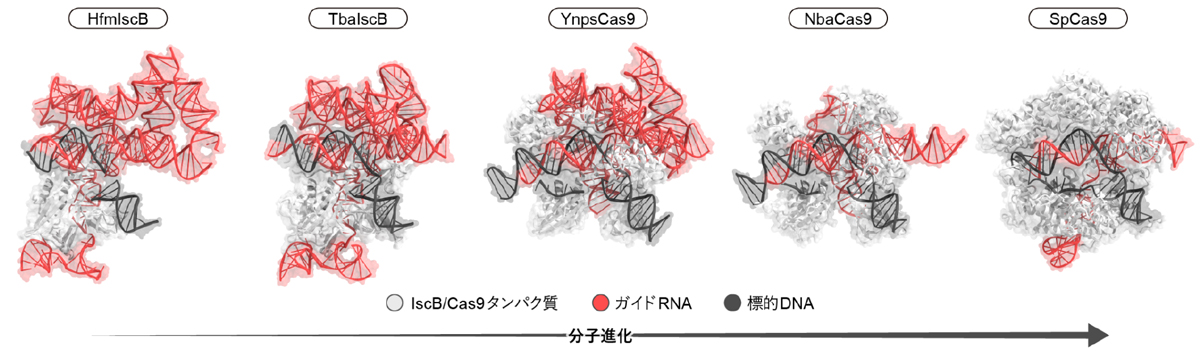
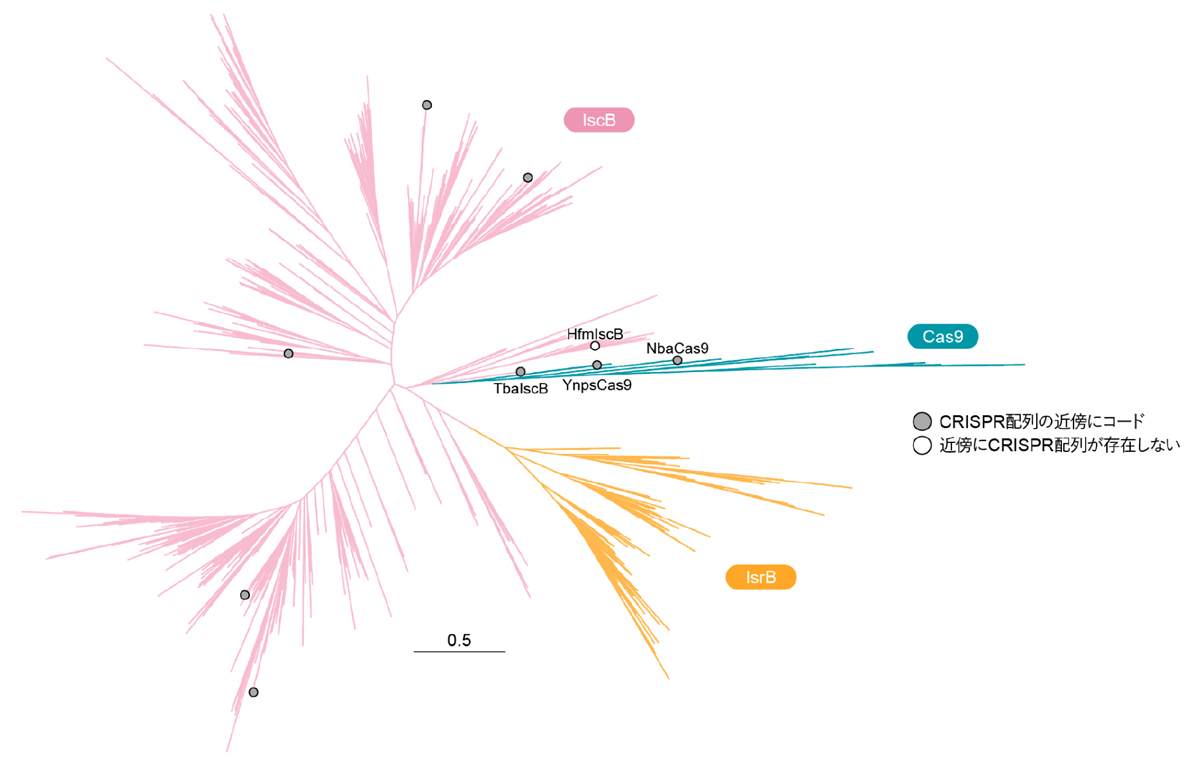
まず、複数のIsrB、IscB、および、Cas9のアミノ酸配列を用いて系統解析を行いました(図2)。cas9遺伝子の近傍にはCRISPR配列がコードされている一方で、通常のiscB遺伝子およびisrB遺伝子の近傍にはCRISPR配列は存在しません。ヒト糞便メタゲノム(注9)由来のHfmIscBをコードするiscB遺伝子の近傍にはCRISPR配列はコードされていない一方、一部のiscB遺伝子の近傍にはCRISPR配列が存在していました。その中でも、Tissierellia綱細菌に由来するTbaIscBはCas9と系統的に近いことから、IscBからCas9への進化中間体に似ていることが示唆されました。また、イエローストーン湖メタゲノムに由来するYnpsCas9とNitrospirae門細菌に由来するNbaCas9は系統的にIscBに近いことから、これらが進化的に初期のCas9に似ていることが示唆されました。
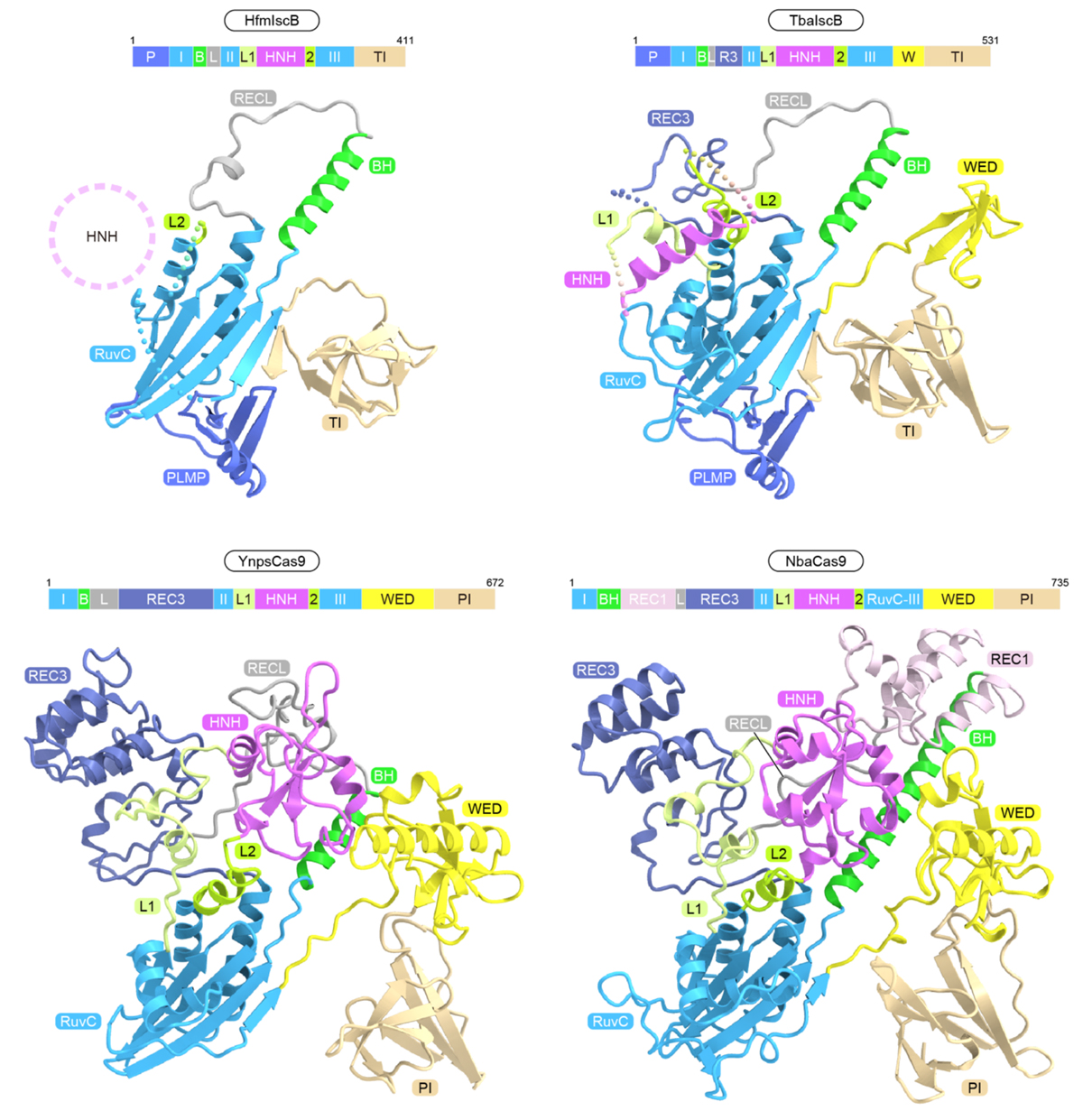
次に、HfmIscB、TbaIscB、YnpsCas9、NbaCas9に関して、クライオ電子顕微鏡を用いて、タンパク質(IscBまたはCas9)、ガイドRNA、および、標的DNAからなる複合体の立体構造を決定しました(図3)。IscBタンパク質およびCas9タンパク質はガイドRNAと複合体を形成し、ガイド配列は標的DNAとヘテロ二本鎖を形成していました。HfmIscBとTbaIscBはRuvCおよびHNHヌクレアーゼドメインに加えて、PLMPドメイン、BH(bridge helix)、REC(recognition)リンカー、TI(TAM-interacting)ドメインを持っていました(図4)。BHとRECリンカーはRNA–DNAヘテロ二本鎖と相互作用し、標的DNA認識に関与していました。HfmIscBとは異なり、TbaIscBはREC3ドメインを持っており、REC3ドメインはRNA–DNAヘテロ二本鎖と相互作用していました。YnpsCas9とNbaCas9はRuvCおよびHNHドメイン、BH、RECリンカー、WED(wedge)ドメイン、PI(PAM-interacting)ドメインを持つ一方、PLMPドメインは持っていませんでした(図4)。さらに、Cas9は多数のαヘリックスから構成されるREC3ドメインやREC1ドメインを持ち、RNA–DNAヘテロ二本鎖を認識していました。REC3ドメインを持つTbaIscB、YnpsCas9、NbaCas9はそれぞれ19、21、21塩基対のRNA–DNAヘテロ二本鎖を認識する一方、REC3ドメインを持たないHfmIscBは16塩基対の短いRNA–DNAヘテロ二本鎖を認識します。これらの立体構造の違いと生化学解析の結果から、IscBからCas9への分子進化の過程において、REC3ドメインの獲得によって、認識できるRNA–DNAヘテロ二本鎖の長さが伸長し、DNA切断の特異性が向上したことが示唆されました。
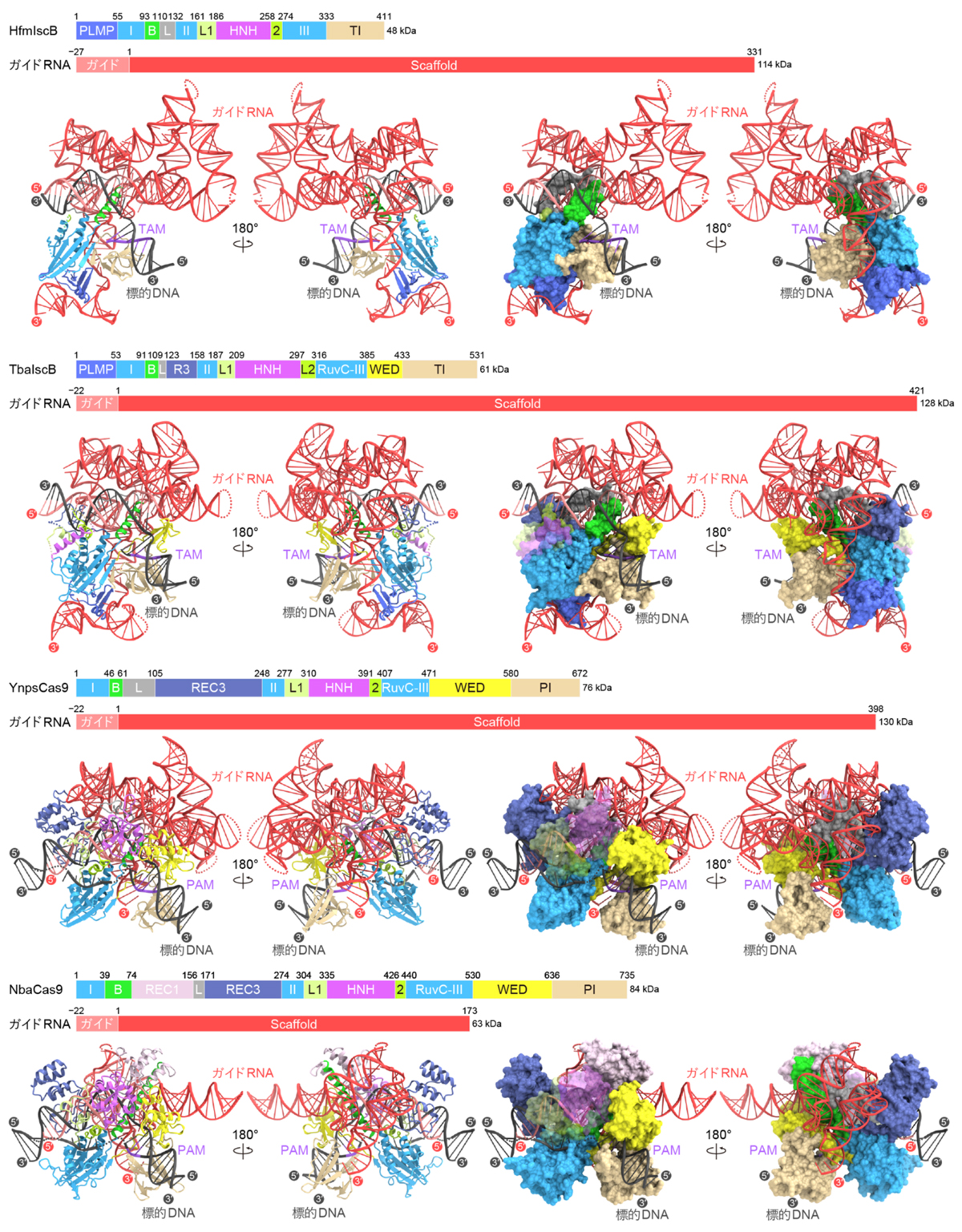
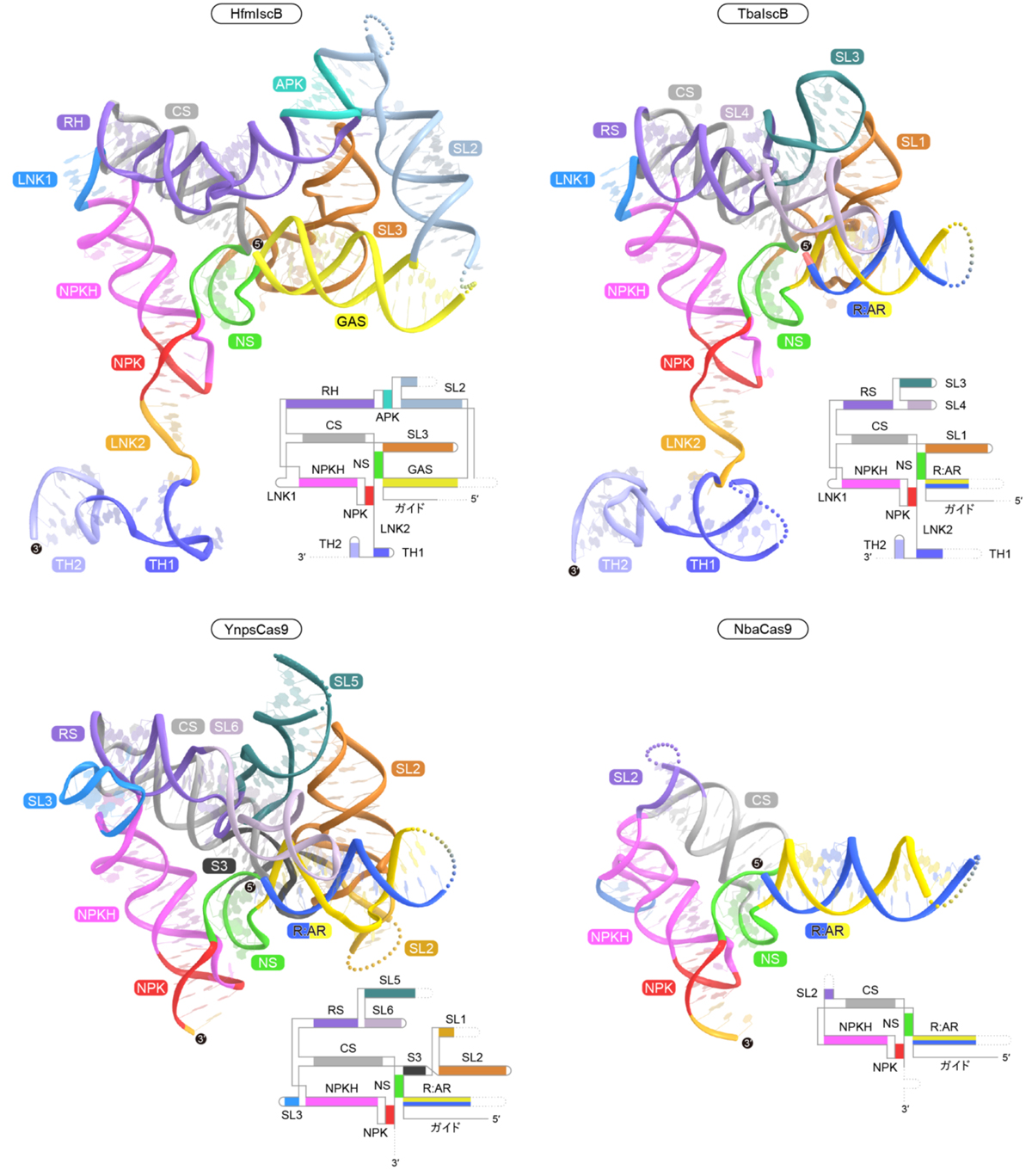
ガイドRNAはガイド配列に加えて、IscB/Cas9タンパク質との結合やDNA認識に関わる足場から構成されます(図5)。HfmIscBとTbaIscBのガイドRNAはTH(terminal hairpin)を持ち、THはPLMPドメインやRuvCドメインと相互作用することで、IscB–ガイドRNA複合体を安定化していました。それに対し、Cas9がPLMPドメインを持たないことと一致して、YnpsCas9とNbaCas9のガイドRNAはTHを欠失していました。CRISPR配列の近傍にコードされるTbaIscBのガイドRNAは、Cas9のガイドRNAと配列と構造が似ていることから、IscBからCas9へ進化の過程において、1本のガイドRNAが2本に分割されたことが示唆されました。
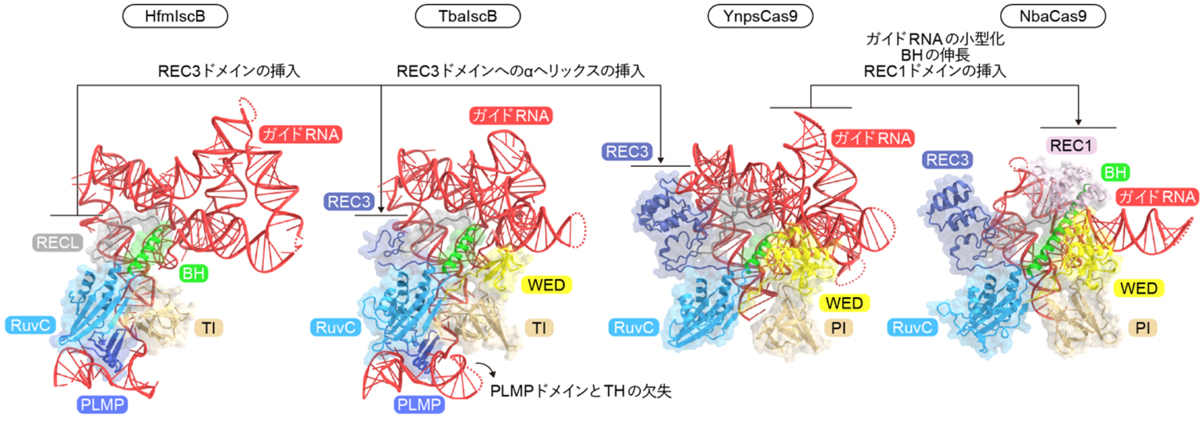
さらに、4つの立体構造を比較することで、IscB/Cas9タンパク質とそれらのガイドRNAが共進化してきたことが明らかになりました(図6)。HfmIscBとTbaIscBがPLMPドメインとTHを持つ一方、YnpsCas9とNbaCas9はその両方を欠失しています。生化学解析により、TbaIscBはガイドRNAからTHを欠失させてもDNA切断活性を示すことが明らかになりました。したがって、IscBからCas9への進化において、ガイドRNAからTHが欠失したのち、PLMPドメインが欠失したことが示唆されました。また、HfmIscB、TbaIscB、および、YnpsCas9は短いBHとRECリンカーを持ち、ガイドRNAは標的DNA認識に関わる大きな足場領域を形成しています。それに対して、NbaCas9は長いBHとREC1ドメインを持ち、ガイドRNAは小型化しています。したがって、Cas9の進化初期において、標的DNA認識に関わるガイドRNAの要素が、構造的および機能的にREC1ドメインに置き換わったことが示唆されました。
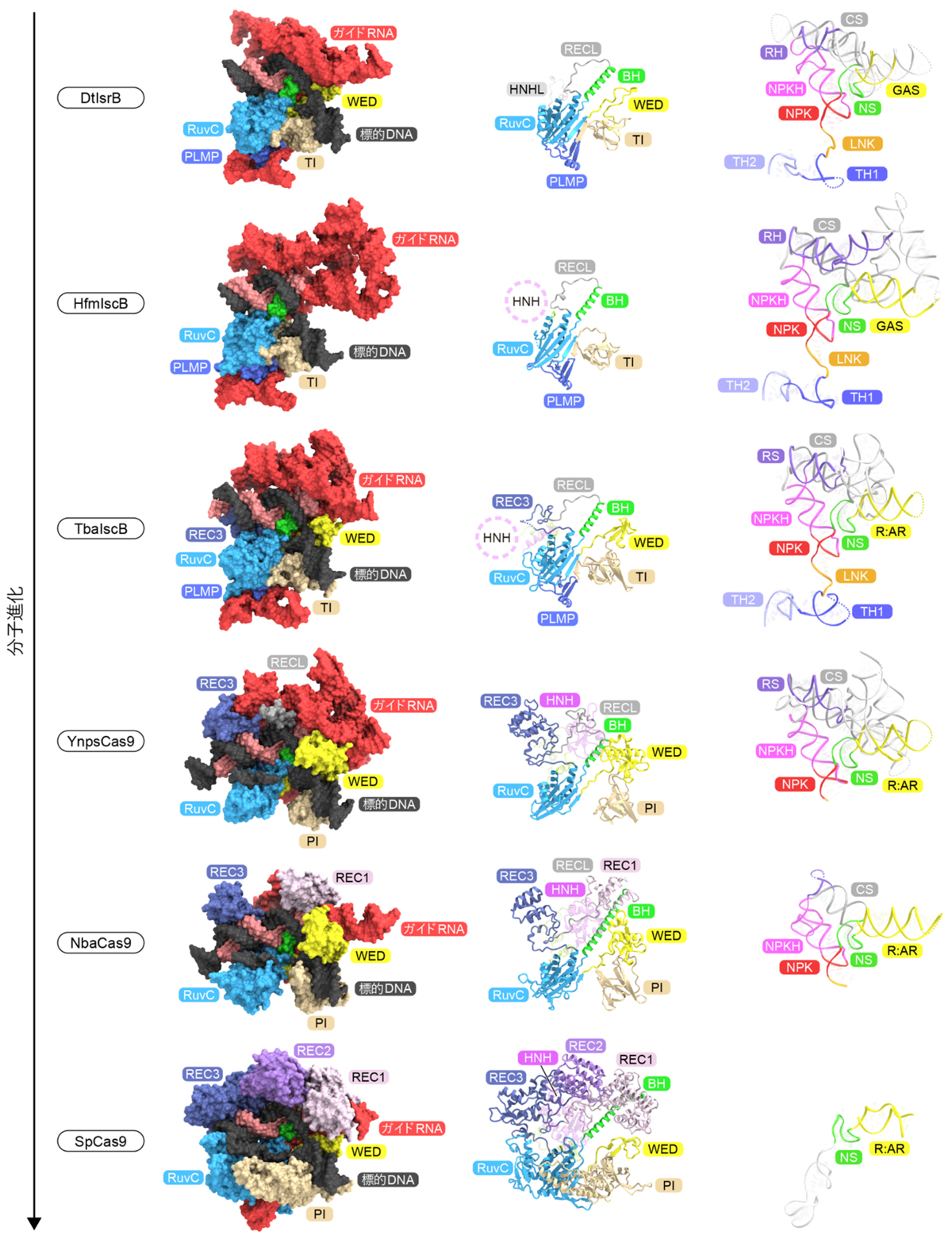
最後に、今回決定した4つのIscBおよびCas9の立体構造を、過去に決定されたDesulfovirgula thermocuniculi由来のIsrB(DtIsrB)とStreptococcus pyogenes由来のCas9(SpCas9)の立体構造と比較しました(図7)。先行研究から、IsrBにHNHヌクレアーゼドメインが融合しIscBが生じたことがわかっていました。本研究では、IscBから初期のCas9への進化において鍵となるステップを明らかにしました。(1)まずIscBがCRISPR配列の近傍にコードされ、(2)REC3ドメインを獲得し、(3)ガイドRNAのTHとPLMPドメインを欠失することでCas9へ進化しました。次に、(4)Cas9がBHを伸長しREC1ドメインを獲得しました。さらに、小型のCas9はREC2ドメインの獲得を伴うタンパク質の大型化、および、ガイドRNAの小型化によってSpCas9のような高いDNA切断活性を持つCas9へと進化しました。
発表者・研究者等情報
東京大学
大学院工学系研究科
長畑 直人 博士課程
山田 崇太 博士課程
平泉 将浩 助教
先端科学技術研究センター
岡崎 早恵 学術専門職員
山下 恵太郎 准教授
西増 弘志 教授
兼:東京大学大学院工学系研究科 教授
東京科学大学総合研究院
諫山 縁 特任研究員
加藤 一希 准教授
Broad Institute of MIT and Harvard
Soumya Kannan Postdoctoral Fellow
Feng Zhang Professor
National Institutes of Health
Eugene V. Koonin NIH Distinguished Investigator
論文情報
- 雑誌名:
- 「Nature Structural & Molecular Biology」(オンライン版:1月30日)
- 題 名:
- Structural visualization of the molecular evolution of CRISPR-Cas9
- 著者名:
- Naoto Nagahata†, Kazuki Kato†, Sota Yamada†, Soumya Kannan, Sae Okazaki, Yukari Isayama, Masahiro Hiraizumi, Keitaro Yamashita, Eugene V. Koonin, Feng Zhang, Hiroshi Nishimasu*(†筆頭著者、*責任著者)
- DOI:
- 10.1038/s41594-025-01743-x
- URL:
- https://doi.org/10.1038/s41594-025-01743-x
研究助成
本研究は、国立研究開発法人日本医療研究開発機構(AMED)「創薬等先端技術支援プラットフォーム(BINDS)(課題番号:JP21am0101115)」、科研費「特別研究員奨励費(課題番号:25KJ1023)」、「基盤研究(S)(課題番号:25H00436)」、「学術変革領域研究(A)(課題番号:21H05281)」、科学技術振興機構(JST)戦略的創造研究推進事業「CREST(課題番号:JPMJCR23B6)」、三菱財団「自然科学研究助成」、稲盛財団「InaRISフェローシップ」、武田科学振興財団「武田報彰医学研究助成」、「ライフサイエンス研究助成」などの支援により実施されました。
用語解説
- (注1)CRISPR-Cas獲得免疫機構
原核生物のもつ生体防御機構のひとつで、原核生物に感染するウイルス由来の核酸(DNAやRNA)の分解を担う。 - (注2)ガイドRNA
Cas9やIscBなどのヌクレアーゼと複合体を形成し、ガイド配列と相補的な標的DNAにヌクレアーゼを誘導する。Cas9のガイドRNAはCRISPR配列とその周辺から産生される。IscBとIsrBのガイドRNAはトランスポゾンの末端から産生される。 - (注3)トランスポゾン
ゲノム上を移動できるDNA配列。動く遺伝子とも呼ばれる。 - (注4)ゲノム編集技術
生命の設計図であるゲノムDNAの塩基配列を改変する技術。CRISPR-Cas9を利用することにより効率的なゲノム編集が可能になった。 - (注5)クライオ電子顕微鏡
液体窒素冷却下でタンパク質などの分子に電子線を照射し、試料の観察を行うための装置。タンパク質や核酸の立体構造の決定に利用されている。 - (注6)ヌクレアーゼドメイン
核酸を切断する機能をもつ、タンパク質のひとかたまりの領域。 - (注7)PAM/TAM配列
PAM(protospacer adjacent motif)とTAM(transposon/target adjacent motif)は、それぞれCas9とIscB/IsrBが認識する標的DNA上の塩基配列。ガイド配列と相補的な標的DNA配列の下流に存在する。 - (注8)系統解析
アミノ酸配列や塩基配列を比較することで、生物や遺伝子の進化的な関係を推定する手法。多くの場合、系統関係は木の形をした系統樹で表される。 - (注9)メタゲノム
環境中(海水、土壌、腸内など)の試料から直接収集されたゲノムDNAの情報。培養を行わずにDNA配列が決定されるため、培養困難な微生物を含む多様な微生物のゲノムDNAの情報が含まれる。
問合せ先
東京大学先端科学技術研究センター 構造生命科学分野
教授 西増 弘志(にします ひろし)
関連タグ
