

ナノサイズの凝縮体が成長せず安定に存在する機構を解明
―鎖長非対称性が生む「エントロピー的電荷分離」が液滴合体を抑制―
- 研究成果
2026年6月4日
東京大学
発表のポイント
- 液液相分離で生じた液滴は、通常は合体やオストワルド熟成によって時間とともに成長すると考えられてきましたが、ナノサイズの生体凝縮体は長時間ほぼ一定の大きさを保つことがあります。本研究は、その謎を説明する新しい物理機構を明らかにしました。
- 反対電荷をもつ高分子の鎖長の非対称性により、全体としては電気的に中性でも、界面でエントロピー的電荷分離が起こって液滴が正味の電荷を帯び、液滴同士の間に長距離の静電反発が生じることを示しました。
- 実験・理論・流体粒子動力学シミュレーションを組み合わせることで、古典的なブラウン運動誘起合体と、今回見いだした合体律速粗大化を統一的に記述できることを示し、生体凝縮体や人工ソフトマターに共通する安定化原理を与えました。
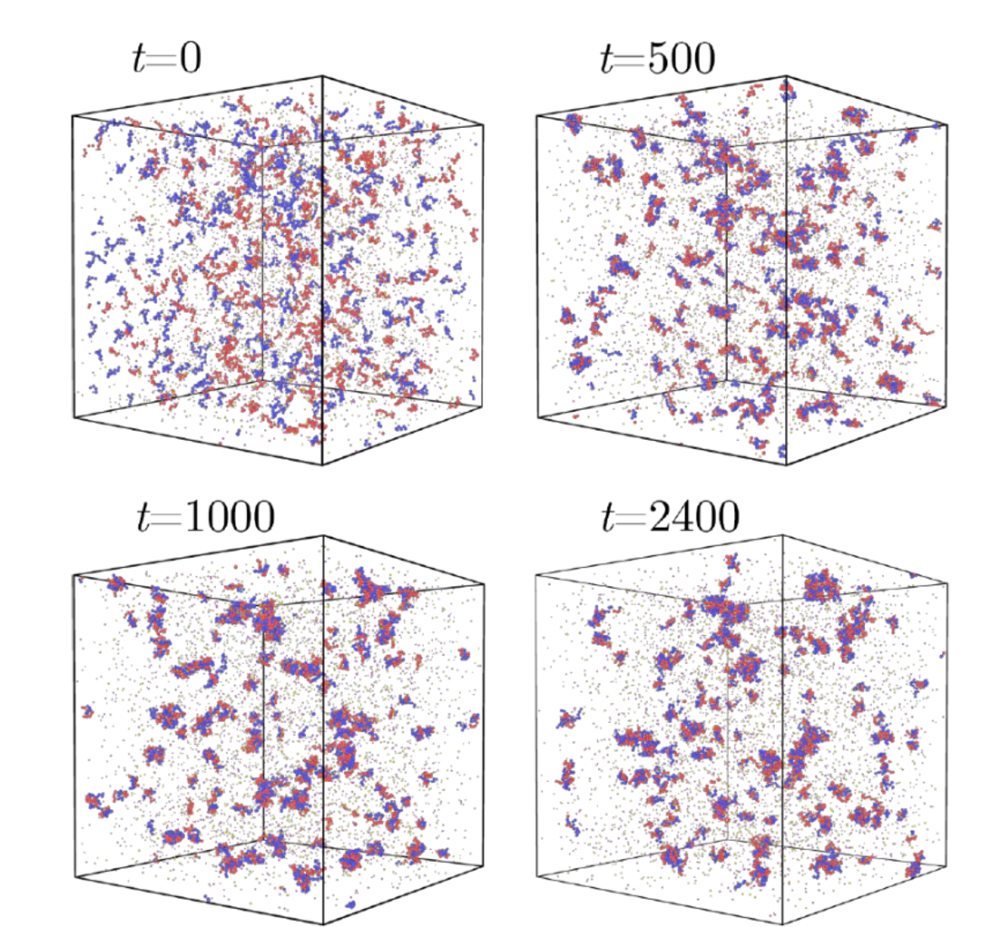
発表概要
東京大学先端科学技術研究センターの田中肇特任研究員(同大名誉教授)らは、香港大学、香港科技大学(広州)、ジョンズ・ホプキンス大学、香港城市大学などとの国際共同研究により、液液相分離(注1)において、ナノサイズの凝縮体が長時間にわたりほとんど成長せず安定に存在できる一般的な物理機構を明らかにしました。
細胞内の生体凝縮体(注2)や、高分子からなるコアセルベート(注3)の液滴は、液滴状でありながら、しばしばナノサイズのまま長時間安定に存在します。しかし古典的な液液相分離理論では、液滴はブラウン運動による衝突合体(注4)やオストワルド熟成(注5)によって連続的に成長すると予想され、このようなサイズ安定性の起源はよく分かっていませんでした。
本研究では、反対電荷をもつ高分子からなるコアセルベートをモデル系として用い、低濃度条件で形成されたナノ液滴が、12時間以上にわたってほとんど成長しないことを見いだしました(図1参照)。さらに、その原因が、高分子鎖長の非対称性(注6)に起因する界面でのエントロピー的電荷分離(注7)にあることを明らかにしました。短い高分子鎖は配置エントロピー(注8)を稼ぐため希薄相へ出やすく、その結果、液滴界面には未中和の電荷が残り、液滴同士の静電反発が合体を抑えます。
この成果は、細胞内で観測される生体凝縮体のサイズ安定性や、人工ナノ凝縮体の設計原理に新しい視点を与えるものであり、ソフトマター、生命科学、ナノテクノロジーにまたがる広い応用が期待されます。
本成果は2026年6月3日(米国東部時間)米国物理学会誌「Physical Review Letters」に掲載されました。また、本論文は注目度の高い研究として、Editors’ Suggestion に選出されました。

ー研究者からのひとことー
液滴が小さいまま安定に存在するには、従来は細胞内の能動的な制御や複雑な生化学的機構が必要だと考えられがちでした。今回の研究では、もっと受動的で一般的な物理法則として、高分子の長さの非対称性そのものが、界面で電荷分離を生み、液滴の合体を止め得ることが見えてきました。凝縮体の安定性は、化学組成だけでなく、分子鎖長とエントロピーの協奏によっても決まるということです。
(東京大学先端科学技術研究センター 特任研究員/東京大学名誉教授 田中肇)
発表内容
液液相分離は、均一な溶液が濃厚相と希薄相に分かれる普遍的現象であり、ソフトマターや細胞内自己組織化の理解において中心的な概念です。特に近年、タンパク質やRNAが形成する生体凝縮体は、遺伝子発現、シグナル伝達、ストレス応答など多様な生命機能に関わることが明らかになってきました。一方で、こうした凝縮体の多くがナノサイズのまま長時間安定に存在することは、従来の相分離理論では十分に説明できない未解決問題でした。古典理論では、液滴はブラウン運動による衝突合体やオストワルド熟成によって成長を続けると考えられてきたためです。
これまで、このサイズ安定性の原因として、ATP(注9)消費を伴う能動的制御、クロマチン(注10)や細胞骨格による物理的拘束、界面に吸着する分子による安定化などが提案されてきました。しかし、近年では、こうした複雑な細胞内環境がなくても、比較的単純な分子系でナノ凝縮体が長く安定に存在することが報告されており、より一般的な物理機構の存在が示唆されていました。
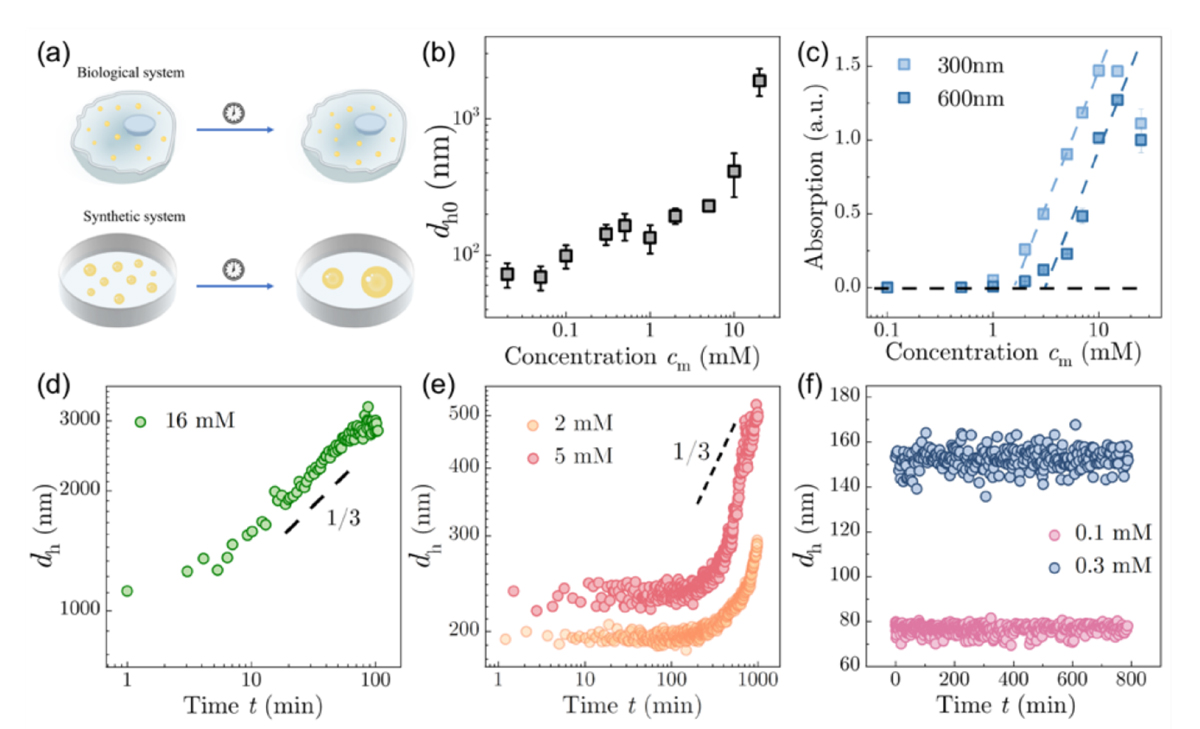
本研究では、負電荷をもつ高分子と正電荷をもつ高分子からなるコアセルベート系を用いて、液滴サイズの時間発展を、動的光散乱(注11)で詳細に測定しました。その結果、高濃度ではミクロンスケール液滴が時間の1/3乗則に従って成長し、古典的なブラウン運動誘起合体の予測と整合する一方、低濃度ではナノサイズ液滴が12時間以上にわたりほとんど成長しないことが分かりました(図1参照)。さらに中間濃度では、しばらく安定に見えた後に急成長する遅延成長領域も観測され、濃度に応じて三つの異なるコアシニング領域が存在することが示されました。
研究チームはこれを説明するために、液滴同士が拡散によって出会う速度だけでなく、出会った後に実際に合体できる速度が液滴サイズに依存する理論モデルを構築しました。その結果、小さな液滴では合体速度が著しく低下し、古典的な、べき乗則成長(注12)ではなく、合体に律速された極めて遅い成長、すなわち、合体律速粗大化(注13)が現れることを示しました。この理論は、低濃度・中間濃度・高濃度の各領域で観測された実験結果を定量的によく再現しました。
では、なぜ小さな液滴ほど合体しにくいのでしょうか。ゼータ電位(注14)測定の結果、コアセルベート液滴は顕著な表面電荷を持ち、その絶対値は液滴サイズとともに減少することが分かりました。また、塩を加えて静電相互作用を遮蔽すると、液滴の初期サイズは大きくなり、高塩濃度では古典的な1/3乗則成長が回復しました。これらの結果は、液滴間の静電反発がナノ液滴の長期安定性を支えていることを強く示しています。
さらに重要なのは、この表面電荷が、全体として中性の条件でも自発的に生じる点です。本研究では、二つの高分子間の鎖長非対称性に注目しました。短い高分子鎖は、濃厚相にとどまるよりも希薄相へ出るほうが配置エントロピーの点で有利であるため、界面近傍で正負電荷の分布に偏りが生じます。つまり、化学量論的(注15)には中性であっても、界面ではエントロピー的電荷分離が起こり、液滴は正味の電荷を帯びるのです。
この機構をさらに検証するため、研究チームは長距離静電相互作用と流体力学相互作用を取り入れた流体粒子動力学シミュレーション(注16)を行いました。その結果、鎖長非対称性がある場合にのみ、界面で未中和電荷が現れ、ナノ液滴の正味電荷密度がサイズとともに減少すること、また対称な鎖長の場合には急速な古典的成長に戻ることが確認されました。すなわち、鎖長非対称性そのものが合体律速粗大化機構を作動させる本質的要因であることが理論・数値計算の両面から示されました。
本研究は、凝縮体のサイズ安定性を、特定の生体成分や能動的制御に依存しない一般的な受動的原理として理解できることを示した点に大きな意義があります。特に、比較的低濃度の条件でエントロピー的電荷分離が最大化されやすいという結果は、細胞内でしばしば見られる亜飽和条件下の凝縮体形成とも整合的です。今後、本成果は、生体凝縮体の形成・安定化機構の理解に加え、人工細胞、ドラッグデリバリー、機能性ソフトマテリアル、ナノ粒子集合体の設計にも新しい指針を与えると期待されます。
発表者・研究者等情報
東京大学先端科学技術研究センター 極小デバイス理工学分野
田中 肇 特任研究員/東京大学名誉教授
香港大学機械工学科
チェン フェイペン
香港科技大学(広州)先進材料学域
ユアン ジアシン
ジョンズ・ホプキンス大学 生物物理学科
チャン ヤオジュン
香港城市大学 生体医工学科
シャム ホーチョン
論文情報
- 雑誌名:
- Physical Review Letters
- 題 名:
- Entropic charge separation as a general mechanism arresting nanoscale condensate coarsening
- 著者名:
- Feipeng Chen, Jiaxing Yuan*, Yaojun Zhang*, Hajime Tanaka*, Ho Cheung Shum* *責任著者
- DOI:
- 10.1103/g1lp-rtd7
研究助成
本研究は、文部科学省科学研究費 特別推進研究(JP20H05619)、香港研究資助局(RGC)、中国国家自然科学基金(NSFC)、ジョンズ・ホプキンス大学スタートアップ基金などの支援を受けて実施されました。
用語解説
- (注1)液液相分離(LLPS)
一様な溶液が、濃厚相と希薄相の二つの液体相に分かれる現象。細胞内ではタンパク質やRNAが局所的に濃縮した生体凝縮体形成の基盤になる。 - (注2)生体凝縮体
細胞内に形成される膜を持たない液滴状集合体。液体的性質をもちつつ、特定分子を濃縮して反応場や制御場として働く。 - (注3)コアセルベート
反対電荷をもつ高分子同士が静電相互作用により会合してできる濃厚液滴相。生体凝縮体の単純化モデルとして広く用いられる。 - (注4)ブラウン運動による衝突合体
液滴が熱揺らぎによるブラウン運動で出会い、衝突・合体しながら成長する古典的コアシニング機構。通常は液滴半径が時間の1/3乗で増大する。 - (注5)オストワルド熟成
小さな液滴が溶けて大きな液滴が成長する現象。界面曲率による化学ポテンシャル差に起因する。 - (注6)鎖長の非対称性
反対電荷をもつ高分子同士で、片方の分子鎖が他方より著しく長い状態。本研究ではこの非対称性が液滴界面での電荷偏りを生む本質的要因となった。 - (注7)エントロピー的電荷分離
全体としては電気的に中性であっても、分子鎖長の違いなどにより、界面で正負電荷の分布に偏りが生じる現象。本研究では短鎖高分子が希薄相へ出やすいことがその起源である。 - (注8)配置エントロピー
配置エントロピーとは、分子や高分子が空間内で取りうる並び方の多さを表す量です。より多くの配置をとれるほど配置エントロピーは大きくなります。本研究では、短い高分子鎖が希薄相でより多くの配置をとれることが、エントロピー的電荷分離の起源になっています。 - (注9)ATP
ATPはアデノシン三リン酸の略称で、細胞内でエネルギーを受け渡す主要な分子です。細胞はATPを利用してさまざまな反応や運動を進めます。生体凝縮体では、ATPを消費する過程がその形成や安定性に影響することがあります。 - (注10)クロマチン
クロマチンとは、細胞核の中でDNAがヒストンなどのタンパク質と結びついてできる複合体です。クロマチンは遺伝情報を収納するだけでなく、DNAの使われやすさや遺伝子発現の制御にも重要な役割を果たします。生体凝縮体の研究では、クロマチンが核内で物理的な障害物や足場として働き、凝縮体の移動や合体、成長を抑える要因になることがあります。 - (注11)動的光散乱
動的光散乱とは、溶液中や分散液中にある微粒子がブラウン運動によって絶えず動くことで、散乱光の強さが時間的に揺らぐ現象を利用して、粒子の大きさを調べる測定法です。一般に、粒子が小さいほど速く動き、大きいほどゆっくり動くため、この揺らぎを解析することで粒子の流体力学的直径を求めることができます。本研究では、コアセルベート液滴のサイズとその時間変化を評価するために用いました。 - (注12)べき乗則成長
べき乗則成長とは、ある量が時間のべき乗に比例して増加する成長のしかたです。例えば、粒子や液滴の大きさ Rが時間 tに対して R∝t^αのように変化する場合、これをべき乗則成長といいます。指数 αの値によって成長の速さや仕組みが異なります。本研究では、高濃度で形成されたコアセルベートが α≈1/3のべき乗則に従って成長し、これは古典的な液滴粗大化の特徴を示しています。 - (注13)合体律速粗大化
液滴同士は拡散で出会うものの、合体そのものに高い障壁があるため、成長全体が合体速度によって律速される粗大化機構。本研究では小さな液滴ほど合体しにくいことが示された。 - (注14)ゼータ電位
分散粒子の表面近傍に形成される電位差を反映する物理量。粒子や液滴がどの程度帯電しているかを示す指標であり、分散安定性と密接に関係する。 - (注15)化学量論的
化学量論的とは、反応や結合に関わる成分どうしの量の比が、化学式や電荷中和の条件に対応してちょうどつり合っていることを指します。本研究では、正電荷と負電荷をもつ高分子の量比が全体としてつり合っている、すなわち電気的に中性となる条件を意味しています。ただし、そのような条件でも、界面では分子の分布に偏りが生じ、局所的には電荷の不均衡が現れることがあります。。 - (注16)流体粒子動力学シミュレーション(FPD)
高分子・液滴・流体の運動を粗視化して扱う数値計算手法。長距離静電相互作用や流体力学相互作用を取り込みながら、相分離や凝縮体の長時間ダイナミクスを解析できます。
問合せ先
東京大学名誉教授
東京大学先端科学技術研究センター
特任研究員 田中 肇(たなか はじめ)
