

微生物Xの献身
―細胞死を誘導する新規メッセージ物質を発見!―
- プレスリリース
2026年6月25日
東京大学
発表のポイント
- ある種の細菌はIII-D2型CRISPR-Casシステムを用いてウイルスの感染を防いでいるが、そのメカニズムは不明だった。
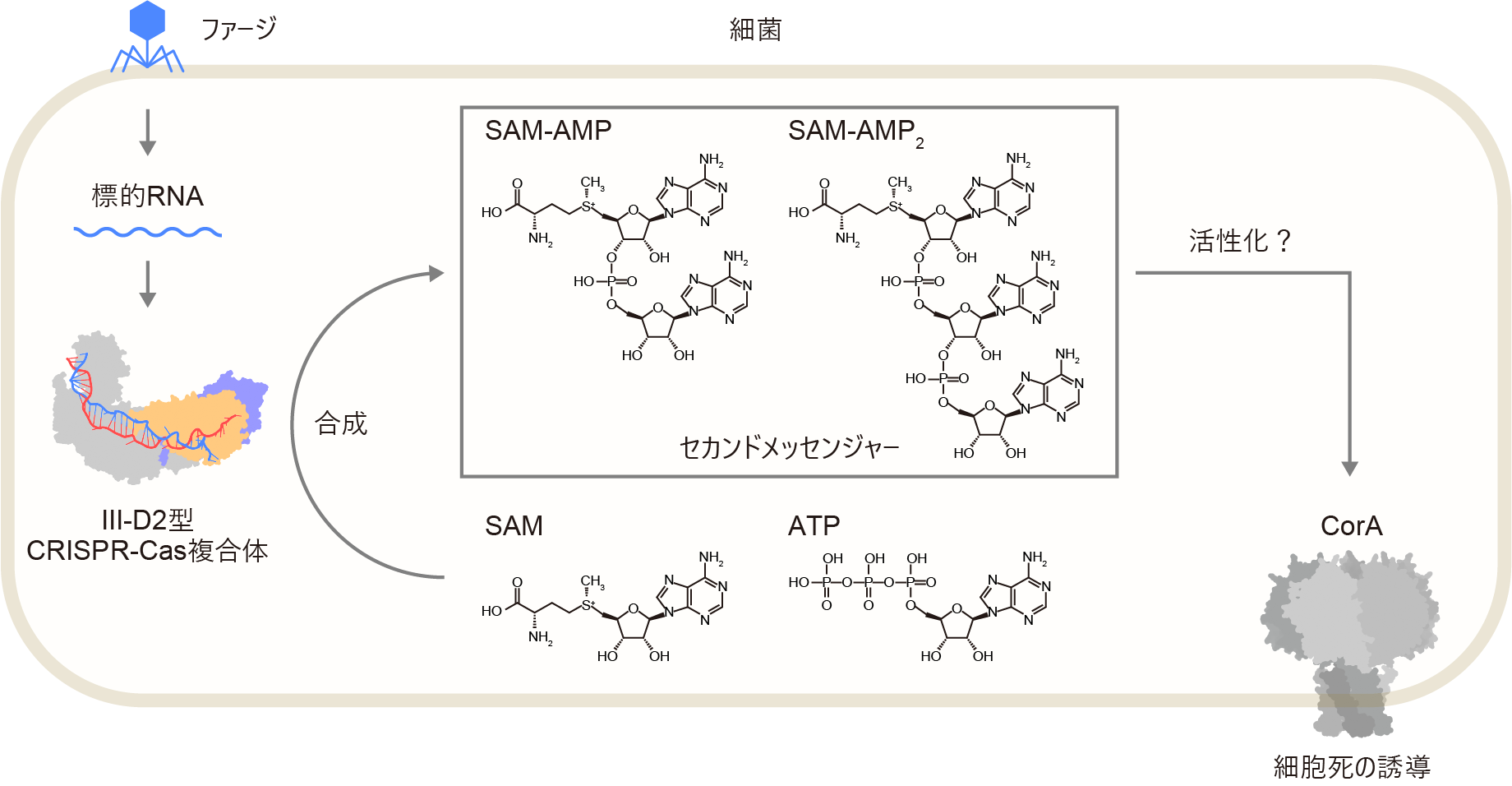
- Cas10、Cas7-5、Cas7x3、および、ガイドRNAからなるIII-D2型CRISPR-Cas複合体は、ウイルスRNAを検知すると、ATPとS-アデノシルメチオニン(SAM)からSAM-AMPとSAM-AMP2を合成することを明らかにした。
- クライオ電子顕微鏡構造解析により、III-D2型CRISPR-Cas複合体がSAM-AMPとSAM-AMP2を合成する分子メカニズムを明らかにした。
- SAM-AMPとSAM-AMP2は膜タンパク質CorAに作用し、細胞死を引き起こすことにより、ウイルス感染から細胞集団を守ることを明らかにした。
概要
原核生物の持つIII型CRISPR-Casシステム(注1)では、Casタンパク質-ガイドRNA複合体がウイルス(ファージ)に由来する外来RNAを認識すると、セカンドメッセンジャー(注2)を合成し、細胞死が引き起こされることにより、ウイルス感染から細胞集団が守られることが知られています。先行研究から、III-A型およびIII-B型CRISPR-Casシステムでは、環状オリゴアデニル酸(cOA)(注3)やSAM-AMP(注4)がセカンドメッセンジャーとしてはたらくことがわかっていましたが、III-D2型CRISPR-Casシステムの機能は不明でした。東京大学大学院工学系研究科化学生命工学専攻の満田義久大学院生、同大学先端科学技術研究センターの西増弘志教授らの研究チームは、同大学先端科学技術研究センターの大澤毅准教授、Harvard Medical SchoolのJonathan S. Gootenberg博士、Omar O. Abudayyeh博士との共同研究として、III-D2型CRISPR-Casシステムによる抗ファージ防御機構を解明しました。まず、III-D2型CRISPR-Casシステムでは、Casタンパク質-ガイドRNA複合体(III-D2型CRISPR-Cas複合体)が標的RNAを認識すると、セカンドメッセンジャーとして、SAM-AMP、および、新規分子であるSAM-AMP2を合成することを明らかにしました。さらに、クライオ電子顕微鏡(注5)を用いて、III-D2型CRISPR-Cas複合体の立体構造を決定し、その合成メカニズムを明らかにしました。さらに、これらのセカンドメッセンジャーが膜タンパク質CorA(注6)に作用し細胞死を誘導することにより、ファージ感染から細菌集団が守られることを明らかにしました。本成果は、2026年6月24日(英国夏時間10:00)に英国科学誌「Nature Communications」オンライン版に掲載されました。
CRISPR-Cas9に代表されるように、外来核酸をガイドRNA依存的に切断するCRISPRシステムが注目されていますが、核酸結合をトリガーとしてシグナル分子を合成するCas10というCasタンパク質も存在します。Cas10が合成するシグナル分子としてこれまで知られていたのは、複数のATPが環状につながったcOAとSAMにATPが結合したSAM-AMPで、CRISPR-Casシステム以外では確認されていない特殊な分子でした。今回の研究で、III-D2型CRISPR-Casシステムでは、SAMに2つのATPが結合したSAM-AMP2が合成されることがわかりました。ファージと原核生物の関係は軍拡競争に例えられることが多く、その影響か、免疫機構に関連するシグナル分子はファージに干渉されないように独創的な構造をしているものが多いです。原核生物がもつ未知の免疫機構には想像もできない新規分子が眠っていると思います。(東京大学大学院工学系研究科 大学院生 満田義久)
実に面白い。(東京大学先端科学技術研究センター 教授 西増弘志)
発表内容
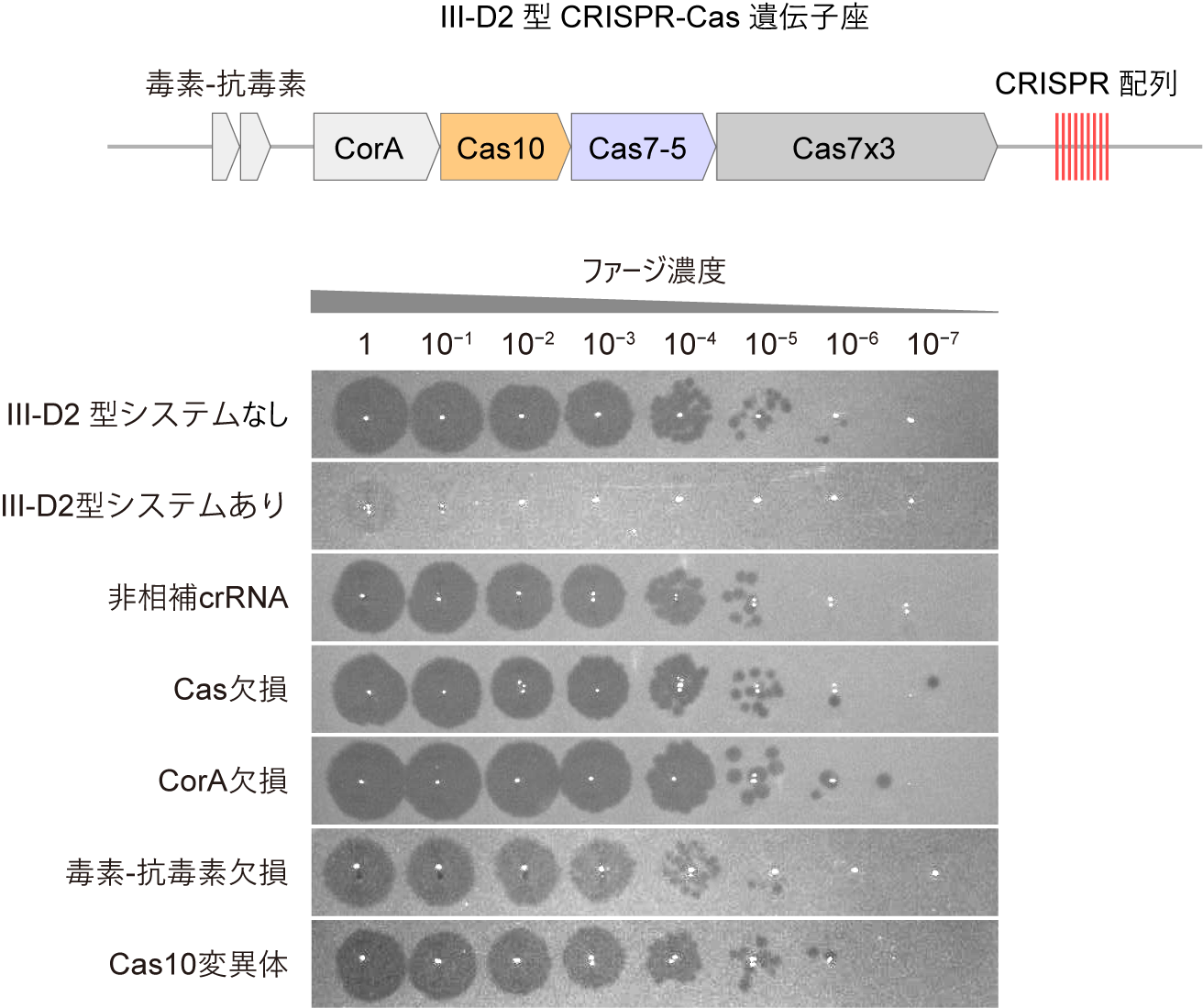
細菌や古細菌などの原核生物は、ウイルス(ファージ)の感染に対抗するため、多様な抗ファージ防御機構を獲得してきました。CRISPR-Casシステムでは、CRISPR RNA(crRNA)がCasタンパク質と複合体を形成し、外来核酸を配列特異的に認識します。なかでも、III-A/B型CRISPR-Casシステムでは、複数のCasタンパク質(Cas5、Cas7、Cas10、Cas11)とcrRNAがIII-A/B型CRISPR-Cas複合体を形成します。III-A/B型複合体が標的RNAに結合すると、Cas10サブユニットが活性化し、ATPを基質として環状オリゴアデニル酸(cOA)と呼ばれるセカンドメッセンジャーが合成されます。cOAは補助エフェクタータンパク質を活性化し、ウイルスに感染した細胞の生育阻害や細胞死を誘導することにより、細菌集団レベルでの感染拡大を防ぎます。CRISPR-Casシステムによって補助エフェクタータンパク質は異なり、RNA分解、DNA分解、代謝物分解など多様な活性を持ちます。さらに、近年、一部のIII-B型CRISPR-Casシステムでは、ATPとSAMを基質としてSAM-AMPと呼ばれる新規のセカンドメッセンジャーが合成されることが報告され、III型CRISPR-Casシステムの多様性が注目されています。III-D2型CRISPR-CasシステムはIII-A型システムから進化したと考えられており、3つのCas7タンパク質が融合したCas7x3、Cas7とCas5が融合したCas7-5、および、Cas10という3つのタンパク質をコードする遺伝子に加え、膜タンパク質CorAと毒素–抗毒素モジュール(注7)から構成されます(図1)。CorAはSAM-AMPを合成するIII-B型CRISPR-Casシステムの補助エフェクタータンパク質として機能することから、III-D2型CRISPR-Casシステムでは、III-D2型複合体が標的RNAを認識するとSAM-AMPが産生され、CorAが活性化することにより細胞死が誘導されると予想されていました。しかし、III-D2型CRISPR-Casシステムの生理機能は不明でした。
III-D2型CRISPR-Casシステムの抗ファージ防御活性を調べるために、ファージRNAと相補的なcrRNAを設計し、III-D2型CRISPR-Casシステムを発現させた大腸菌にファージを感染させたところ、ファージ感染に対して抵抗性を示すことがわかりました(図1)。一方、crRNAがファージRNAと相補性を持たない場合、防御活性は見られませんでした。また、Casタンパク質、CorA、毒素–抗毒素モジュールの遺伝子をそれぞれ欠失させた場合も防御活性は見られませんでした。さらに、Cas10の活性残基の変異によっても防御活性が消失しました。これらの結果から、III-D2型CRISPR-Casシステムでは、(1)crRNAを介したファージRNAの認識、(2)Cas10によるセカンドメッセンジャーの合成、および、(3)CorAと毒素–抗毒素モジュールの存在、が抗ファージ防御活性に必要であることが示唆されました。
III-D2型CRISPR-Casシステムと標的RNAを発現させた大腸菌は生育阻害を示したことから、III-D2型システムが標的RNAを認識して活性化すると細胞毒性を示すことが明らかになりました。さらに、この細胞毒性にはCorAが不可欠である一方、毒素–抗毒素モジュールは必要でなかったことから、セカンドメッセンジャーはCorAに結合して細胞死を誘導し、毒素–抗毒素モジュールは細胞死の誘導とは異なる役割を持つことが示唆されました。
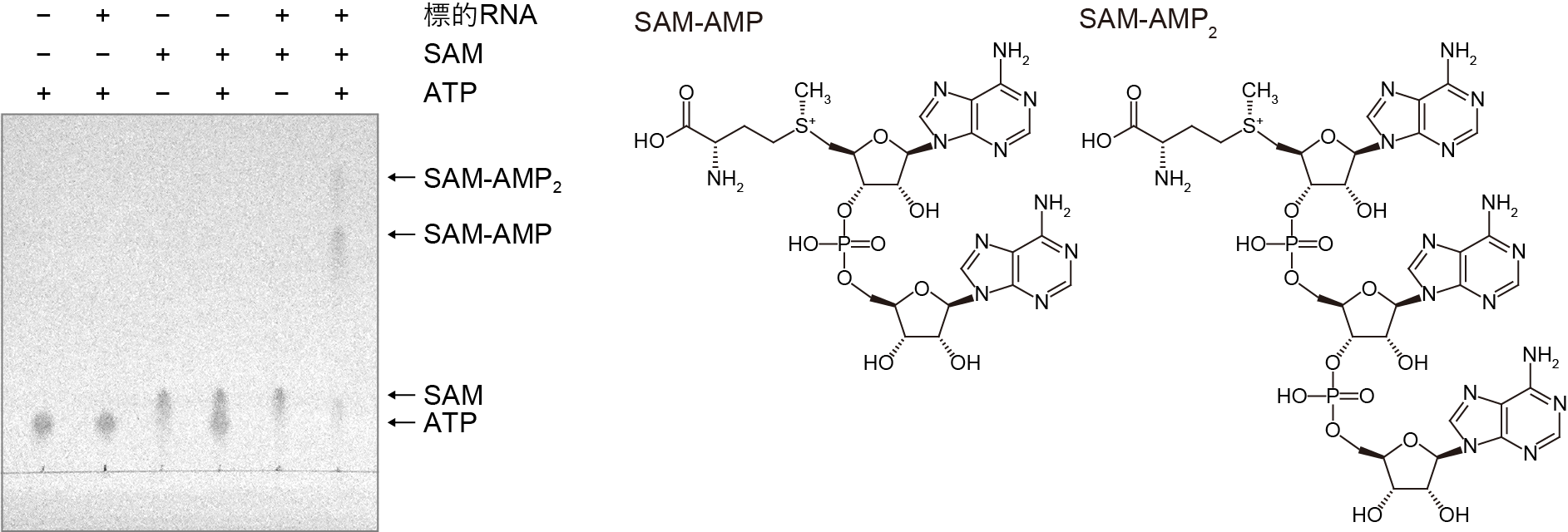
次に、精製したIII-D2型CRISPR-Cas複合体を用いて、セカンドメッセンジャー合成活性を調べました。試験管内でIII-D2型CRISPR-Cas複合体と標的RNA、ATP、SAMを反応させたところ、2種類の生成物が検出されました(図2)。質量分析の結果、1つは既知のセカンドメッセンジャーであるSAM-AMPである一方、もう1つはSAM-AMPにさらにAMPが連結した新規のシグナル伝達分子SAM-AMP2であることが判明しました。注目すべきことに、反応の経過に伴いSAM-AMPが減少し、SAM-AMP2が増加することが明らかになりました。これらの結果から、SAM-AMP2はSAM-AMPとATPから合成されることが示唆されました。
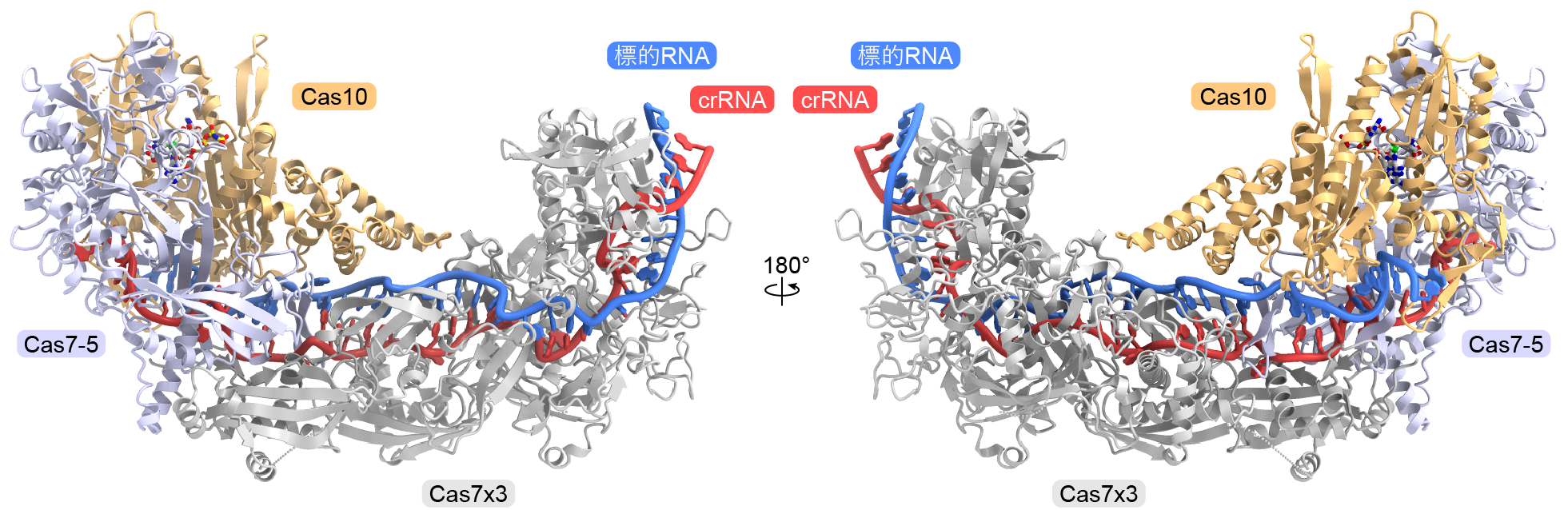
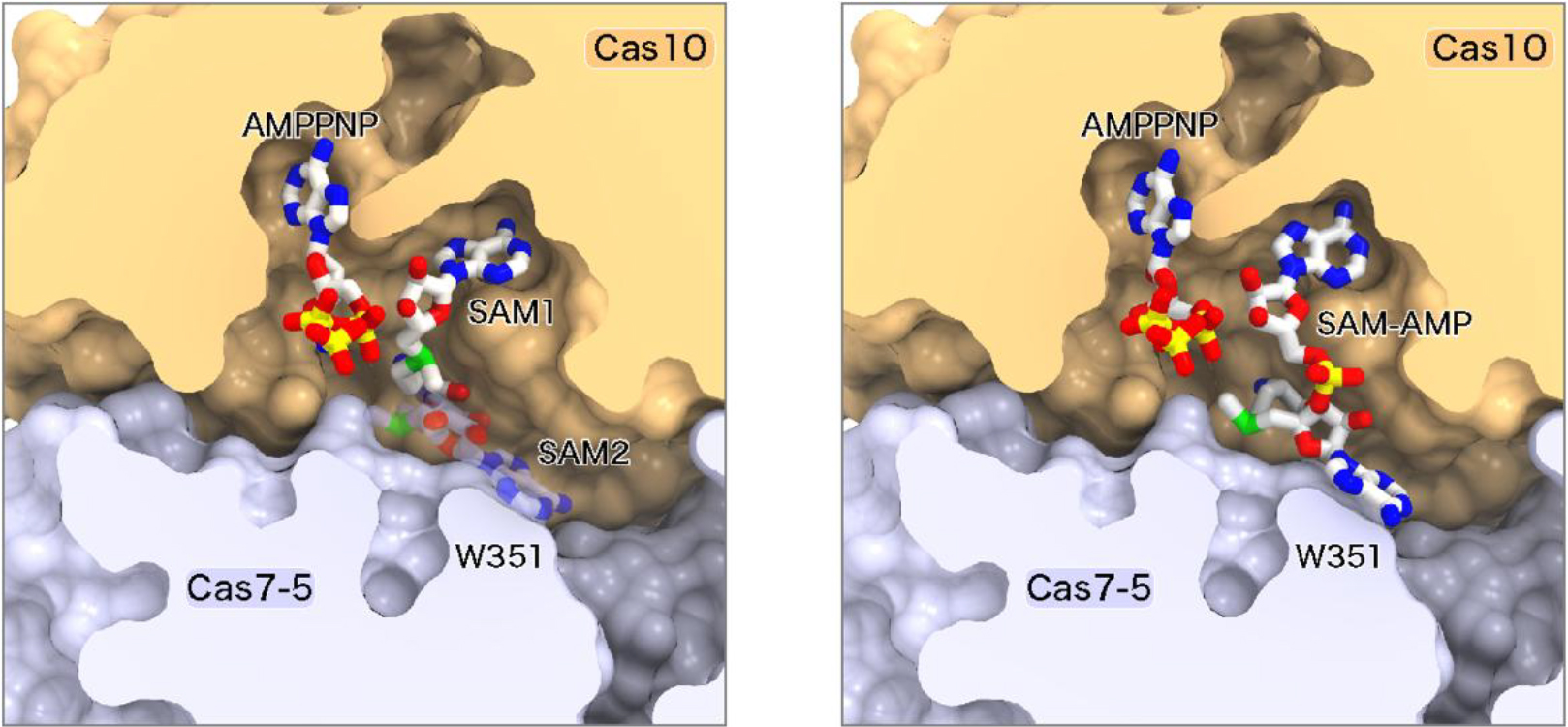
次に、クライオ電子顕微鏡を用いてIII-D2型CRISPR-Cas複合体の立体構造を決定しました(図3)。その結果、Cas10の活性部位にAMPPNP(ATP類似体)とSAMが結合していることが確認されました。さらに、標的RNAの末端領域がCas10に結合すると活性部位がSAM-AMP合成に適した構造に変化することがわかりました。SAMは2つのコンフォメーションをとり、Cas10の活性部位に加えて、Cas7-5のW351残基とも相互作用していました(図4)。W351残基の変異は、SAM-AMP合成活性には影響しなかった一方、SAM-AMP2合成活性を消失させたことから、SAM-AMPがW351残基を介して結合しATPと反応することによりSAM-AMP2が合成されることが示唆されました。
最後に、SAM-AMP合成およびSAM-AMP2合成に関わるアミノ酸残基に変異を導入し、抗ファージ防御活性を調べました。その結果、SAM-AMP2合成のみが損なわれる変異体でも防御活性が低下したことから、SAM-AMPだけでなくSAM-AMP2もIII-D2型CRISPR-Casシステムによる抗ファージ防御に寄与していることが示唆されました。
以上の結果から、III-D2型CRISPR-Casシステムでは、SAM-AMPとSAM-AMP2がシグナル伝達分子としてはたらき抗ファージ防御を実現していることが明らかになりました。SAM-AMPとSAM-AMP2がCorAに作用して細胞死を引き起こすメカニズムや毒素-抗毒素モジュールの役割を解明するには、さらなる研究が必要です。本研究により、CRISPR-Casシステムの理解が深まるとともに、原核生物が合成する新規のセカンドメッセンジャーの多様性が明らかになりました。
発表者・研究者等情報
東京大学
大学院工学系研究科
満田 義久 博士課程
石川 潤一郎 博士課程(研究当時)
長畑 直人 博士課程
平泉 将浩 助教
先端科学技術研究センター
岡崎 早恵 学術専門職員(研究当時)
山下 恵太郎 准教授
大澤 毅 准教授
西増 弘志 教授
兼:東京大学大学院工学系研究科 教授
東京科学大学総合研究院
菅谷 麻希 特任研究員
加藤 一希 准教授
Harvard Medical School
Jonathan S. Gootenberg Investigator
Omar O. Abudayyeh Investigator
論文情報
- 雑誌名:
- 「Nature Communications」(オンライン版:6月24日)
- 題 名:
- Structural mechanism of SAM-AMP and SAM-AMP2 synthesis by the type III-D2 CRISPR effector complex
- 著者名:
- Yoshihisa Mitsuda, Maki Sugaya, Junichiro Ishikawa, Naoto Nagahata, Sae Okazaki, Masahiro Hiraizumi, Kato Kazuki, Jonathan S. Gootenberg, Omar O. Abudayyeh, Tsuyoshi Osawa, Keitaro Yamashita, Hiroshi Nishimasu*(*責任著者)
- DOI:
- https://doi.org/10.1038/s41467-026-74422-0
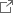
研究助成
本研究は、日本学術振興会(JSPS)科研費「特別研究員奨励費(課題番号:23KJ0720、25KJ1023)」、「若手研究(課題番号:23K14133)」、「基盤研究(S)(課題番号:25H00436)」、科学技術振興機構(JST)戦略的創造研究推進事業「ACT-X(課題番号:JPMJAX232F)」、「CREST(課題番号:JPMJCR23B6)」、国立研究開発法人日本医療研究開発機構(AMED)「創薬等先端技術支援プラットフォーム(BINDS)(課題番号:JP21am0101115)」、AMED(課題番号:23wm0325065)、武田科学振興財団「武田報彰医学研究助成」、稲盛財団InaRISフェローシップなどの支援により実施されました。
用語解説
- (注1)III型CRISPR-Casシステム
原核生物の抗ファージ防御機構であるCRISPR-Casシステムの1つ。Cas5、Cas7、Cas10、Cas11がcrRNAと複合体を形成し、外来RNAを標的とする。Casタンパク質の構成によってIII-A型からIII-I型に分類される。 - (注2)セカンドメッセンジャー
細胞内シグナルを伝達する小分子。III型CRISPR-Casシステムでは、外来RNAを認識するとエフェクター複合体がセカンドメッセンジャーを合成する。セカンドメッセンジャーは補助エフェクタータンパク質に作用し細胞死を誘導する。 - (注3)環状オリゴアデニル酸
複数のATPが環状に繋がった分子。 - (注4)SAM-AMP
S-アデノシルメチオニン(SAM)とAMPからなる分子。SAMはメチル基供与体としても利用される。 - (注5)クライオ電子顕微鏡
液体窒素冷却下でタンパク質などの分子に電子線を照射し、試料の観察を行うための装置。タンパク質や核酸の立体構造の決定に利用されている。 - (注6)CorA
2価イオントランスポーターと配列相同性をもつ膜タンパク質。機能未知ドメインと膜貫通ドメインからなり、SAM-AMPの結合によって活性化し脱分極を起こすと予想されている。 - (注7)毒素–抗毒素モジュール
細胞毒性をもつタンパク質と、毒性を抑えるタンパク質からなる生体システム。原核生物に存在し、環境ストレス応答や免疫応答に関与する。
問合せ先
東京大学先端科学技術研究センター 構造生命科学分野
教授 西増 弘志(にします ひろし)
関連タグ
